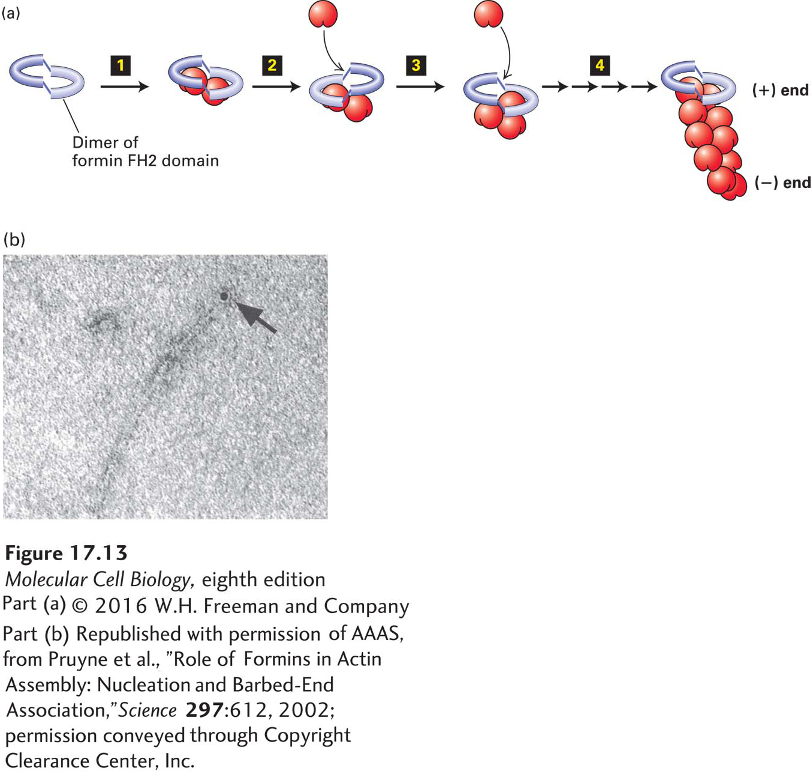
Formins are found in essentially all eukaryotic cells as a diverse family of proteins: seven different classes are present in vertebrates. Although they are diverse, all formin family members have two adjacent domains in common, the so-called FH1 and FH2 domains (formin-homology domains 1 and 2). Two FH2 domains from two individual formin monomers associate to form a doughnut-shaped complex (Figure 17-13a). This complex has the ability to nucleate actin assembly by binding two actin subunits, holding them so that the (+) end of the nascent filament is toward the FH2 domains. The filament can now grow at the (+) end while the FH2 domain dimer remains attached to it. How is this possible? As we saw earlier, an actin filament can be thought of as two intertwined strands of subunits. The FH2 dimer can bind to the two terminal subunits. It then probably rocks between the two end subunits, letting go of one to allow addition of a new subunit and then binding the newly added subunit and freeing up space for the addition of another subunit to the other strand. In this way, rocking between the two subunits on the end, it can remain attached while simultaneously allowing growth at the (+) end (see Figure 17-13a).
[Part (b) republished with permission of AAAS, from Pruyne et al., “Role of Formins in Actin Assembly: Nucleation and Barbed-End Association,” Science297:612, 2002; permission conveyed through Copyright Clearance Center, Inc.]
FIGURE 17-13Actin nucleation by the formin FH2 domain. (a) Formins have a domain called FH2 that can form a dimer and nucleate filament assembly. The dimer binds two actin subunits step 1 and, by rocking back and forth steps 2–4, can allow insertion of additional subunits between the FH2 domain and the (+) end of the growing filament. The FH2 domain protects the (+) end from being capped by capping proteins. (b) The FH2 domain of a formin was labeled with colloidal gold (black dot) and used to nucleate assembly of an actin filament. The resulting filament was visualized by electron microscopy after staining with uranyl acetate. Formins assemble long unbranched filaments.
[Part (b) republished with permission of AAAS, from Pruyne et al., “Role of Formins in Actin Assembly: Nucleation and Barbed-End Association,” Science297:612, 2002; permission conveyed through Copyright Clearance Center, Inc.]
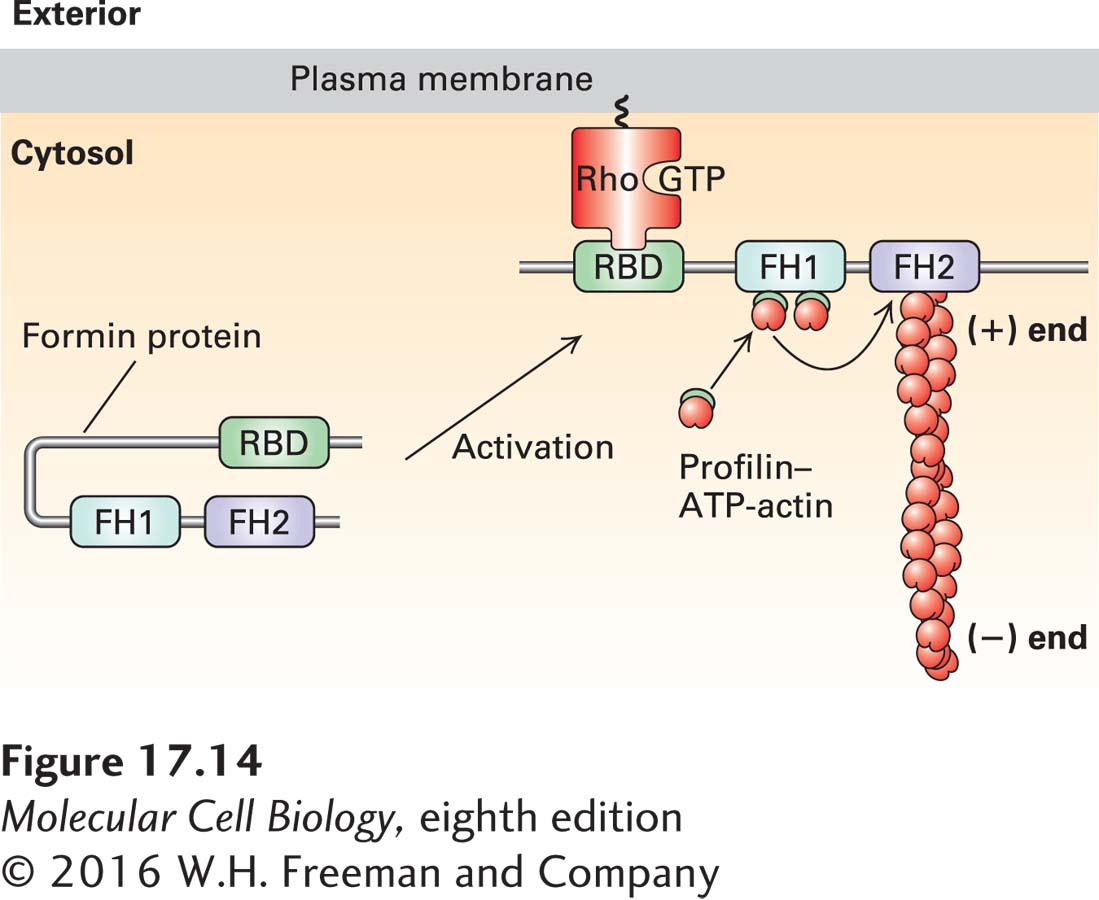
The FH1 domain adjacent to the FH2 domain also makes an important contribution to actin filament growth (Figure 17-14). This domain is rich in proline residues, which serve as sites for the binding of several profilin molecules. We discussed earlier how profilin can exchange the ADP nucleotide on G-actin for ATP to generate profilin–ATP-actin. The FH1 domain behaves as a landing site for profilin–ATP–G-actin to increase the local concentration of these complexes. The actin from these profilin-actin complexes is then fed into the FH2 domain to add actin to the (+) end of the filament, and the profilin is released (see Figure 17-14). Since the formin allows rapid addition of actin subunits to the (+) end, long filaments with a formin at their (+) end are generated (Figure 17-13b). In this manner, formins not only nucleate actin assembly, but have the remarkable ability to remain bound to the (+) end while also allowing rapid assembly there. To ensure the continued growth of the filament, formins bind to the (+) end in such a way that precludes binding of a (+) end capping protein such as CapZ, which would normally terminate assembly.
FIGURE 17-14Regulation of formin activity by an intramolecular interaction. Some of the formin classes found in vertebrates are regulated by an intramolecular interaction. In its inactive state, the formin folds back on itself to inhibit the activity of the FH2 domain. The inactive formin is activated when its Rho-binding domain (RBD) binds to membrane-bound active Rho-GTP, resulting in exposure of the formin’s FH2 domain, which can then nucleate the assembly of a new actin filament. All formins have an FH1 domain adjacent to the FH2 domain; the proline-rich FH1 domain is a site for recruitment of profilin–ATP–G-actin complexes that can then be “fed” into the growing (+) end. For simplicity of representation, a single formin protein is shown, but as shown in Figure 17-13, the FH2 domain functions as a dimer to nucleate actin assembly. Regulation of the Rho family of small GTPases is detailed in Figures 17-41 and 17-43.
To be useful to a cell, formin activity must be regulated. Many formins exist in a folded, inactive conformation as a result of an interaction between the first half of the protein and its C-terminal tail. These formins are activated by membrane-bound Rho-GTP, a Ras-related small GTPase (discussed in Section 17.7). When Rho is switched from its inactive Rho-GDP form to its activated membrane-bound Rho-GTP state, it can bind and activate these formins (see Figure 17-14).
Recent studies have shown that formins are responsible for the assembly of long actin filaments such as those found in muscle cells, stress fibers, filopodia, and the contractile ring that forms during cytokinesis (see Figure 17-4). The actin-nucleating role of formins was discovered only recently, so the roles performed by this diverse protein family are only now being uncovered. Since there are many different formin classes in animals, it is likely that formins will be found to assemble additional actin-based structures.