The Arp2/3 Complex Nucleates Branched Filament Assembly
The Arp2/3 complex is a protein machine consisting of seven subunits, two of which are actin-related proteins (Arp), explaining its name (Figure 17-15a). It is found in essentially all eukaryotes, including plants, yeasts, and animal cells. The Arp2/3 complex alone is a very poor nucleator. To nucleate the assembly of branched actin, Arp2/3 needs to be activated by interacting with a nucleation promoting factor (NPF), in addition to associating with the side of a preexisting actin filament. Although there are many different NPFs, the major NPF family is characterized by the presence of a region called WCA (WH2, connector, acidic). Experiments have shown that if the WCA domain is added to an actin assembly assay together with preformed actin filaments, Arp2/3 becomes a potent nucleator of further actin assembly.
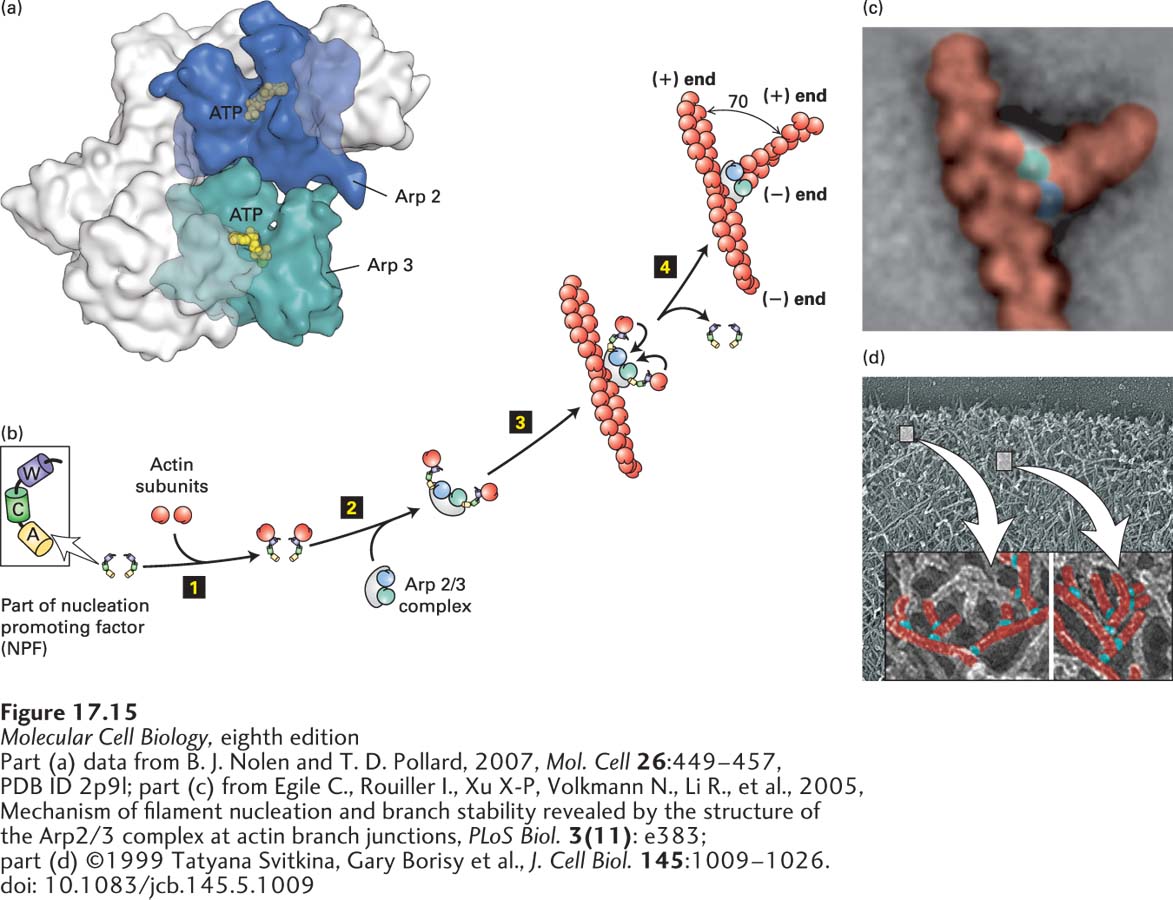
FIGURE 17-15Actin nucleation by the Arp2/3 complex. (a) X-ray crystallographic structure of the Arp2/3 complex, with five of the subunits in gray and the Arp2 and Arp3 subunits in green and blue. (b) To nucleate actin assembly efficiently, Arp2/3 must interact with the activating part of an NPF, shown here with its W (WH2), C (connector), and A (acidic) domains. The first step 1 involves binding of an actin subunit to the W domain of each NPF. Two NPF-actin complexes then bind the Arp2/3 complex step 2. This interaction induces a conformational change in the Arp2/3 complex. After binding of the Arp2/3 complex to the side of an actin filament, the actin subunits delivered by the W domains bind to the Arp2/3 complex step 3, which then initiates the assembly of an actin filament at the available (+) end step 4. The Arp2/3 branch makes a characteristic 70° angle between the old and new filaments. (c) Averaged image compiled from several electron micrographs of Arp2/3 at an actin branch. (d) Image of actin filaments in the leading edge of a motile cell, with a magnification and coloring of individual branched filaments.
How do the Arp2/3 complex and NPFs nucleate the assembly of actin filaments? Two NPFs each bind an actin subunit at their WH2 domains, and together, they activate the Arp2/3 complex through its interaction with their connector and acidic domains. In the inactive Arp2/3 complex, the two actin-related polypeptides—Arp2 and Arp3—are in the wrong configuration to nucleate filament assembly (see Figure 17-15b, step 2). When activated by the NPFs, Arp2 and Arp3 move into the correct configuration, and the complex binds the side of a preexisting actin filament. The actin subunits brought in by the WH2 domains of the NPFs binds to the Arp2/3 template to nucleate filament assembly at the (+) end (Figure 17-15b). The NPFs are released, and the new (+) end then grows as long as ATP–G-actin is available or until it is capped by a (+) end capping protein such as CapZ. The angle between the old filament and the new one is 70° (Figure 17-15c). This is also the angle observed experimentally in branched filaments at the leading edges of motile cells, which are believed to be formed by the action of the activated Arp2/3 complex (Figure 17-15d). As we discuss in subsequent sections, the Arp2/3 complex can be used to drive actin polymerization to power intracellular motility.
Page 788
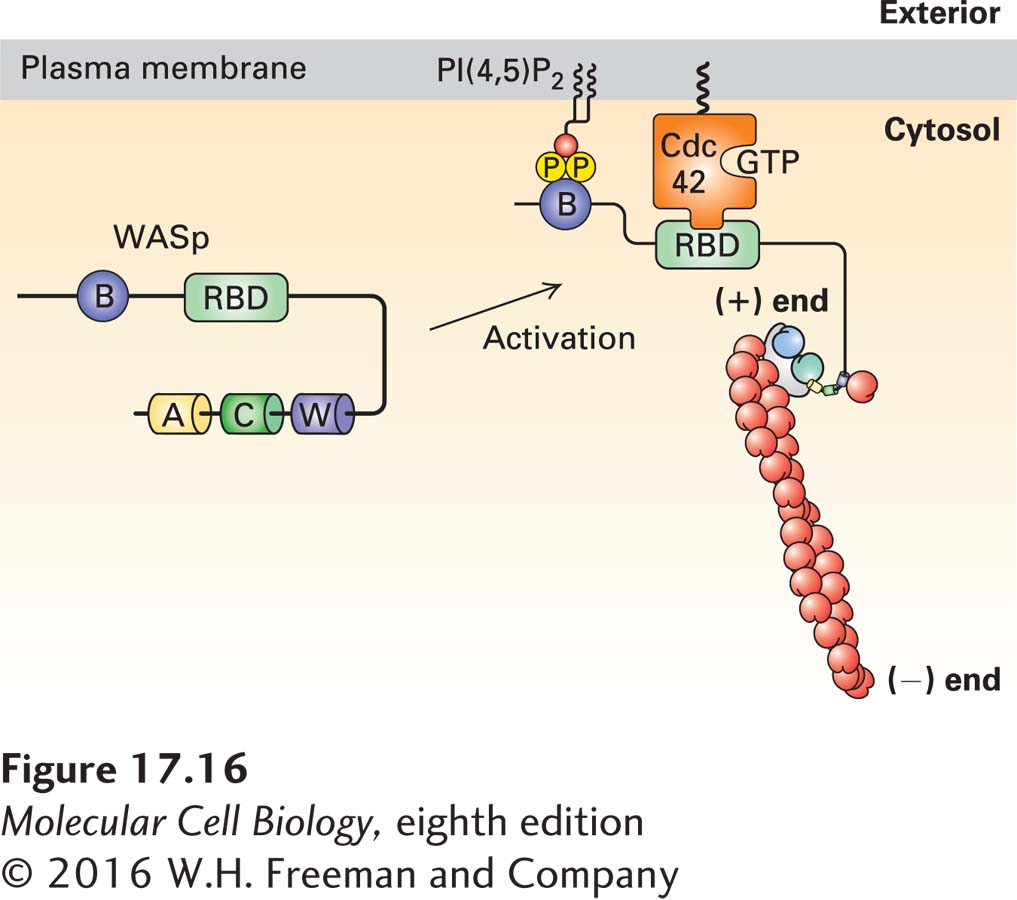
Actin nucleation by the Arp2/3 complex is finely controlled, and the NPFs are part of that regulatory process. One NPF is called WASp, as it is defective in patients with Wiskott-Aldrich syndrome, an X-linked disease characterized by eczema, low platelet count, and immune deficiency. WASp exists in a folded inactive conformation that makes the WCA domain unavailable (Figure 17-16). It is important that this protein be activated only at the plasma membrane, and its activation requires two signals. One signal is the presence of the regulatory phospholipid PI(4,5)P2, which is characteristically enriched in the plasma membrane (Chapter 16). WASp binds PI(4,5)P2 through its basic domain. The second signal is binding of the activated form of the small GTP-binding protein Cdc42, which is itself activated in response to signaling pathways (discussed in Section 17.7). This type of two-signal input, called coincidence detection, ensures that the protein is activated only at the right place—at the plasma membrane—and by the right signaling pathway. Once bound to the two input signals, WASp is opened, and the WCA domain becomes accessible.
Another important NPF is a large protein complex called WAVE, which also has a WCA domain that can activate the Arp2/3 complex. WAVE is also activated by binding of acidic phospholipids and by the active form of another small GTP-binding protein, Rac1. As we discuss in Section 17.7, activation of the Arp2/3 complex by Cdc42 through WASp and by Rac1 through WAVE induces the formation of different microfilament-based structures.
FIGURE 17-16Regulation of the Arp2/3 complex by WASp and PI(4,5)P2. The NPF WASp is inactive due to an intramolecular interaction that masks the WCA domain. It is activated by a coincidence detection mechanism: it must bind both the regulatory phospholipid PI(4,5)P2 through its basic domain (B) and the membrane-bound active small G protein Cdc42-GTP (a member of the Rho family) through its Rho-binding domain (RBD). When activated in this way, the intramolecular interaction in WASp is relieved, allowing the W domain to bind actin and the acidic A domain to activate the Arp2/3 complex. For simplicity, only a single NFP-Arp2/3 interaction is shown. Regulation of the Rho family of small GTPases is detailed in Figures 17-41 and 17-43.
Page 789
Although formins and the Arp2/3 complex are found in fungi, plants, and animals, additional actin nucleators have recently been discovered in animal cells. One of these, called Spire, has four tandem WH2 domains, so it can bind four actin monomers. It does this in a manner that allows the actins to assemble into a filament, although the detailed mechanism remains to be understood. Given that actin filaments perform so many functions in cells, it is not surprising that additional NPF proteins and actin nucleators have recently been described.