Intracellular Movements Can Be Powered by Actin Polymerization
How can actin polymerization be harnessed to do work? As we have seen, actin polymerization involves the hydrolysis of actin-ATP to actin-ADP, which allows actin to grow preferentially at the (+) end and disassemble at the (−) end. If an actin filament were to become fixed in the network of the cytoskeleton and you could bind to and ride on the assembling (+) end, you would be transported across the cell. This is just what the intracellular bacterial parasite Listeria monocytogenes does to get around the cell. The study of Listeria motility was, in fact, the way the nucleating activity of the Arp2/3 protein was discovered. As we will see shortly, Listeria has hijacked a normal cell motility process for its own purposes. We discuss Listeria first, as it is currently much better understood than the normal processes that employ similar mechanisms.
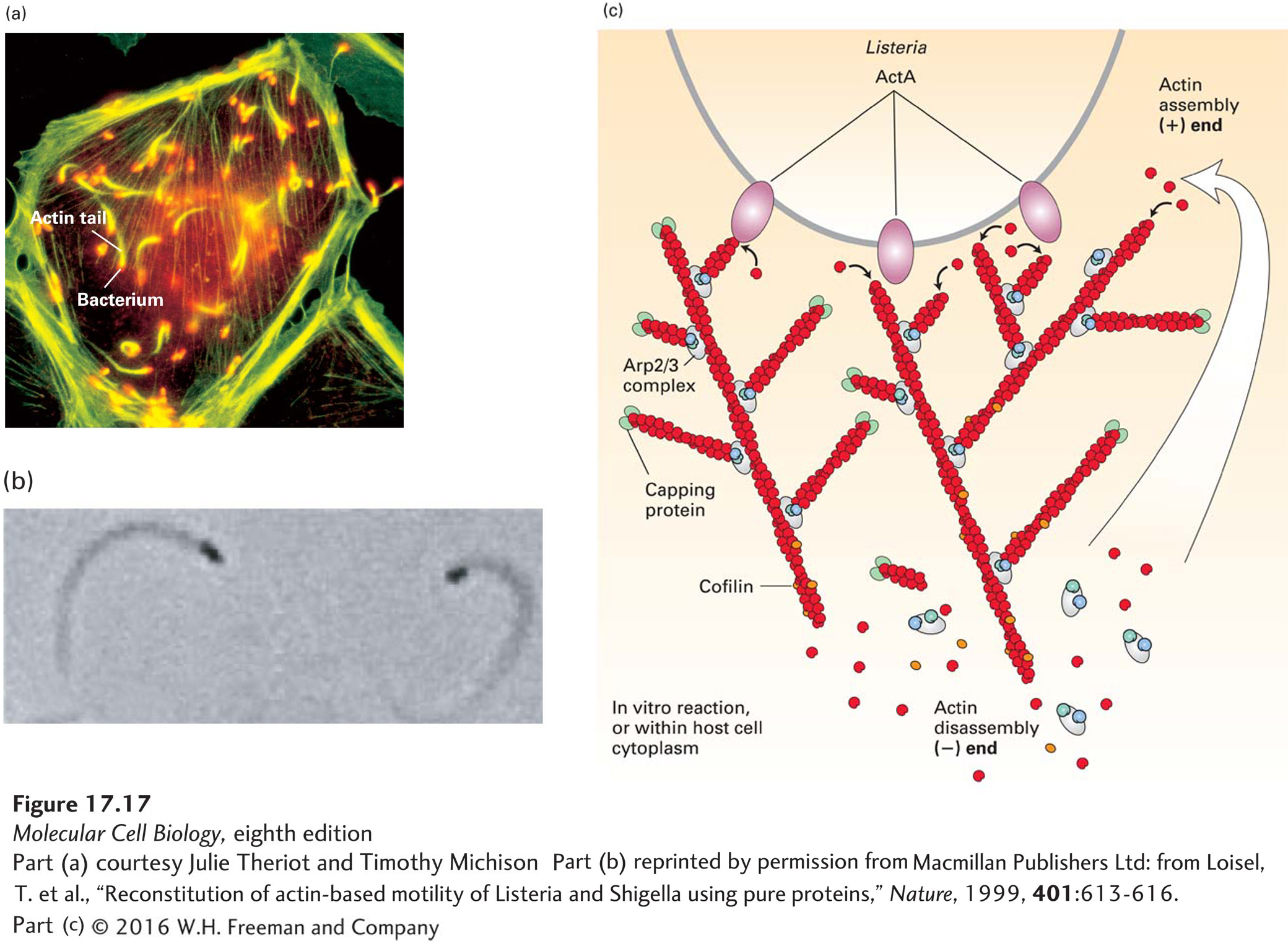
Listeria is a food-borne pathogen that causes mild gastrointestinal symptoms in most adults but can be fatal to elderly or immunocompromised individuals. It enters animal cells and divides in the cytoplasm. To move from one host cell to another, it moves around the cell by polymerizing actin into a comet tail like the plume behind a rocket (Figure 17-17a, b), and when it runs into the plasma membrane, it pushes its way into the adjacent cell to infect it. To do this, it needs to direct the assembly of host-cell actin locally at its back and at the same time confine assembly there so that it efficiently pushes the bacterium forward. How does it do it? Listeria has on its surface a protein called ActA, which mimics an NPF by having an actin-binding site and an acidic region that efficiently activates the Arp2/3 complex (Figure 17-17c). The ActA protein also binds a host protein known as VASP, which has three important properties. First, VASP has a proline-rich region that can bind profilin–ATP-actin and thus enhance ATP-actin assembly into the newly formed barbed ends generated by the Arp2/3 complex. Second, it can hold onto the end of the newly formed filament. Third, it can protect the (+) end of the growing filament from capping by CapZ. These properties allow VASP to enhance actin assembly and confine it to the rear of the bacterium. The assembling filaments then push on the bacterium. Since the filaments are embedded in the stationary cytoskeletal matrix of the cell, the Listeria cell is pushed forward, ahead of the polymerizing actin. Researchers have reconstituted Listeria motility in the test tube using purified proteins to see what the minimal requirements for Listeria motility are. Remarkably, the bacteria will move when just four proteins are added: ATP–G-actin, the Arp2/3 complex, CapZ, and cofilin (see Figure 17-17b, c). We have discussed the role of actin and Arp2/3, but why are CapZ and cofilin needed? As we have seen earlier, CapZ rapidly caps the free (+) end of actin filaments, so when a growing filament no longer contributes to bacterial movement, it is rapidly capped and inhibited from further elongation. In this way, assembly occurs mostly adjacent to the bacterium, where ActA is stimulating the Arp2/3 complex. Cofilin is necessary to accelerate the disassembly of the (−) end of the actin filament, regenerating free actin to keep the polymerization cycle going (see Figure 17-11). This minimal rate of motility can be increased by the presence of other proteins, such as VASP and profilin, as mentioned above.
[Part (a) courtesy Julie Theriot and Timothy Michison; part (b) reprinted by permission from Macmillan Publishers Ltd: from Loisel, T. et al., “Reconstitution of actin-based motility of Listeria and Shigella using pure proteins,” Nature, 1999, 401:613-616.]
EXPERIMENTAL FIGURE 17-17Listeria uses the power of actin polymerization for intracellular movement. (a) Fluorescence microscopy of a cultured cell stained with an antibody to a bacterial surface protein (red) and fluorescent phalloidin to localize F-actin (green). Behind each Listeria bacterium is an actin “comet tail” that propels the bacterium forward by actin polymerization. When the bacterium runs into the plasma membrane, it pushes the membrane out into a structure like a filopodium, which protrudes into a neighboring cell. (b) Listeria motility can be reconstituted in vitro with bacteria and just four proteins: ATP–G-actin, Arp2/3 complex, CapZ, and cofilin. This phase-contrast micrograph shows bacteria (black), behind which are the phase-dense actin tails. (c) A model of how Listeria moves using just four proteins. The ActA protein on the bacterial cell surface activates the Arp2/3 complex to nucleate new filament assembly from preexisting filaments. Filaments grow at their (+) end until capped by CapZ. Actin is recycled through the action of cofilin, which enhances depolymerization at the (−) end of the filaments. In this way, polymerization is confined to the back of the bacterium and propels it forward. Although not essential for motility in vitro, the protein VASP (not shown) binds ActA in vivo to enhance motility, as described in the text.
[Part (a) courtesy Julie Theriot and Timothy Michison; part (b) reprinted by permission from Macmillan Publishers Ltd: from Loisel, T. et al., “Reconstitution of actin-based motility of Listeria and Shigella using pure proteins,” Nature, 1999, 401:613-616.]
Page 790
To move inside cells, the Listeria bacterium, as well as other opportunistic pathogens such as the Shigella species that cause dysentery, hijacks a normal, regulated cellular process involved in cell locomotion. As we discuss in more detail later (Section 17.7), moving cells have a thin sheet of cytoplasm at the front of the cell called the leading edge (see Figures 17-1c, 17-4, and 17-15d). This thin sheet of cytoplasm consists of a dense network of actin filaments that are continually elongating at the front of the cell to push the membrane forward. Factors in the leading-edge membrane activate the Arp2/3 complex to nucleate these filaments. Thus the power of actin assembly pushes the membrane forward to contribute to cell locomotion.