Myosins Have Head, Neck, and Tail Domains with Distinct Functions
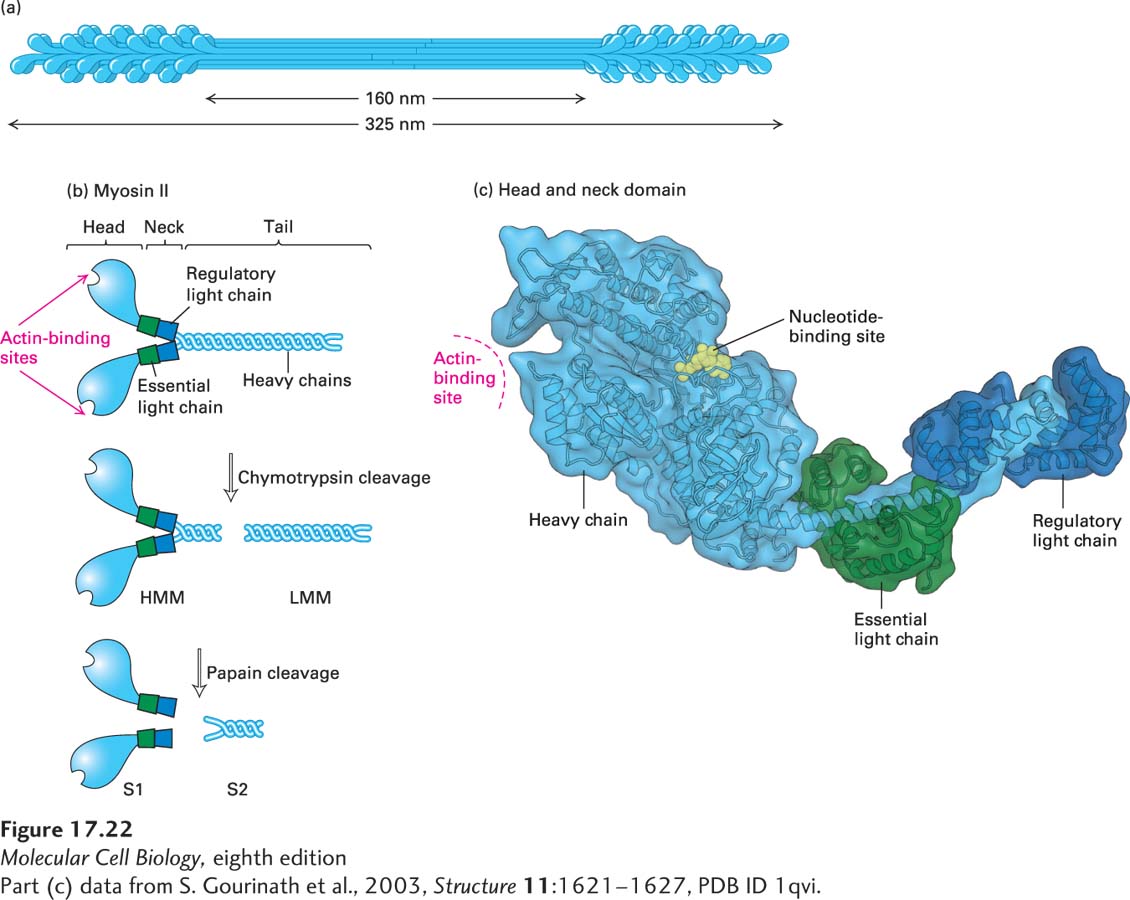
Much of what we know about myosins comes from studies of myosin II isolated from skeletal muscle. In skeletal muscle, hundreds of individual myosin II molecules are assembled into bipolar bundles called thick filaments (Figure 17-22a). In a later section, we will discuss how these myosin filaments interdigitate with actin filaments to bring about muscle contraction. Here we first investigate the properties of the myosin molecule itself.
[Part (c) data from S. Gourinath et al., 2003, Structure11:1621–1627, PDB ID 1qvi.]
FIGURE 17-22Structure of myosin II. (a) Organization of myosin II in filaments isolated from skeletal muscle. Myosin II assembles into bipolar filaments in which the tails form the shaft of the filament and the heads are exposed at the ends. Treatment of bipolar filaments with high salt concentrations and ATP disassembles the filament into individual myosin II molecules. (b) A myosin II molecule consists of two identical heavy chains (light blue) and four light chains (green and dark blue). The tails of the heavy chains form a coiled coil to dimerize; the neck region of each heavy chain has two light chains associated with it. Limited proteolytic cleavage of myosin II generates tail fragments—LMM and S2—and the S1 motor domain. (c) Three-dimensional model of a single S1 head domain shows that it has a curved, elongated shape and is bisected by a cleft. The nucleotide-binding pocket lies on one side of this cleft, and the actin-binding site lies on the other side near the tip of the head. Wrapped around the shaft of the α-helical neck are two light chains. These chains stiffen the neck so that it can act as a lever arm for the head. Shown here is the ADP-bound conformation.
[Part (c) data from S. Gourinath et al., 2003, Structure11:1621–1627, PDB ID 1qvi.]
It is possible to dissolve the myosin thick filament in a solution of ATP and a high concentration of salt, generating a pool of individual myosin II molecules. The soluble myosin II molecule is actually a protein complex consisting of six polypeptide subunits. Two of the subunits are identical high-molecular-weight polypeptides known as myosin heavy chains. Each consists of a globular head domain and a long tail domain, connected by a flexible neck domain. The tails of the two myosin heavy chains intertwine, so that the head regions are in close proximity. The remaining four subunits of the myosin complex are smaller in size and are known as the light chains. There are two types of light chains, the essential light chain and the regulatory light chain. One light chain of each type associates with the neck region of each heavy chain (Figure 17-22b, top). The myosin heavy chain and the two types of light chains are encoded by three different genes.
Page 798
The soluble myosin II molecule has ATPase activity, reflecting its ability to power movements by hydrolysis of ATP. But which part of the myosin complex is responsible for this activity? A standard approach to identifying functional domains in a protein is to cleave the protein into fragments with specific proteases and then ask which fragments have the activity. Soluble myosin II can be cleaved by gentle treatment with the protease chymotrypsin to yield two fragments, one called heavy meromyosin (HMM; mero means “part of”) and the other, light meromyosin (LMM) (Figure 17-22b, middle). The heavy meromyosin can be further cleaved by the protease papain to yield subfragment 1 (S1) and subfragment 2 (S2) (Figure 17-22b, bottom). By analyzing the properties of the various fragments—S1, S2, and LMM—it was found that the intrinsic ATPase activity of myosin resides in the S1 fragment, as does its F-actin–binding site. Moreover, it was found that the ATPase activity of the S1 fragment is greatly enhanced by the presence of filamentous actin, so that fragment is said to have an actin-activated ATPase activity, which is a hallmark of all myosins. The S1 fragment of myosin II consists of the head and neck domains with associated light chains, whereas the S2 and LMM regions make up the tail domain.
X-ray crystallographic analysis of the head and neck domains revealed the shapes of the subunits, the positions of the light chains, and the locations of the ATP-binding and actin-binding sites (Figure 17-22c). At the base of the myosin head is the α-helical neck, where two light-chain molecules wrap around the heavy chain like C-clamps. In this position, the light chains stiffen the neck region. The actin-binding site is an exposed region at the tip of the head domain; the ATP-binding site is also in the head domain, within a cleft opposite the actin-binding site.
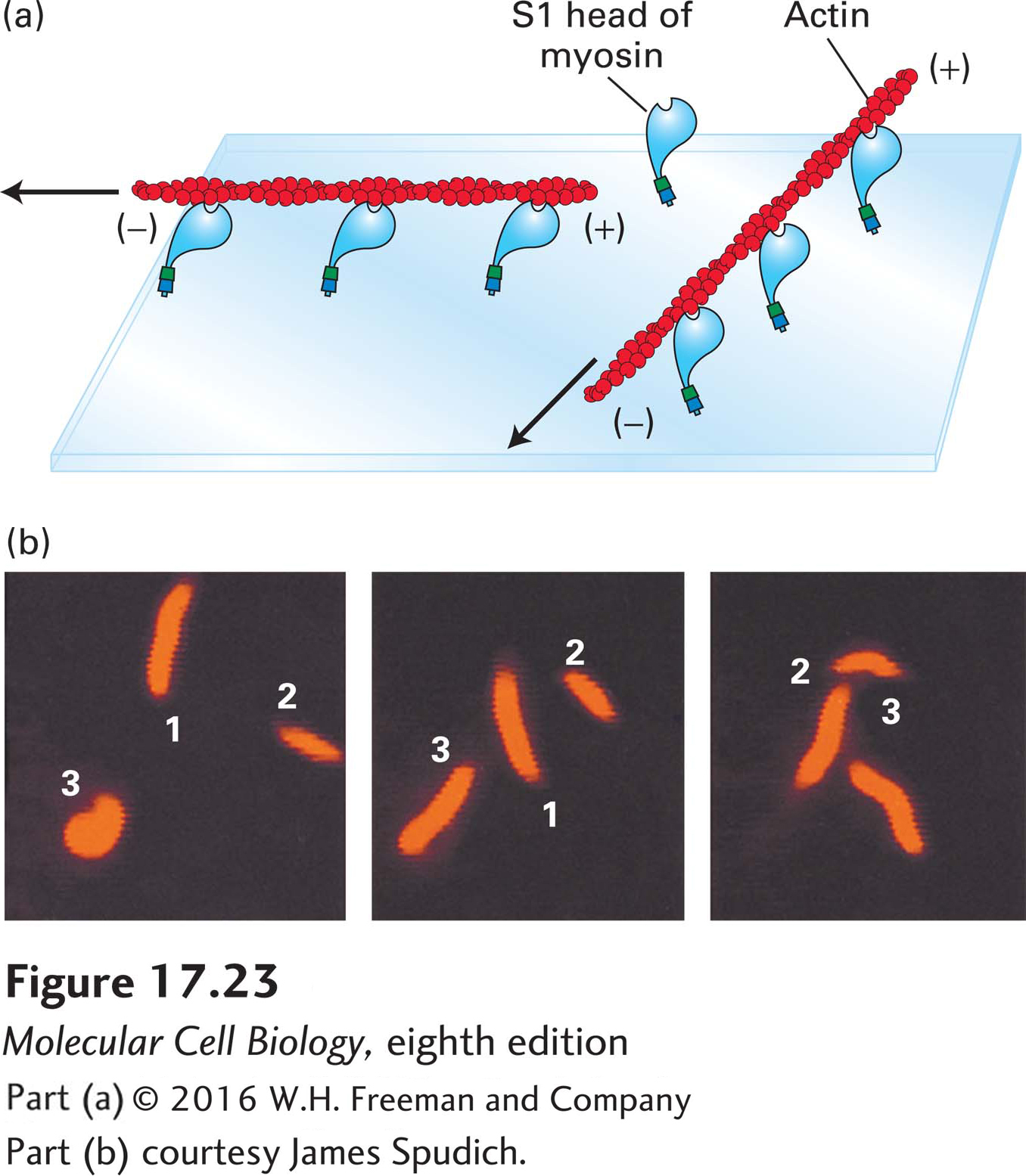
How much of the myosin II molecule is necessary and sufficient for its “motor” activity? To answer this question, one needs a simple in vitro motility assay. In one such assay, the sliding-filament assay, myosin molecules are tethered to a coverslip to which is added stabilized, fluorescently labeled actin filaments. Because the myosin molecules are tethered, they cannot slide; thus any force generated by the interaction of myosin heads with actin filaments forces the filaments to move relative to the myosin (Figure 17-23a). If ATP is present, the added actin filaments can be seen to glide along the surface of the coverslip; if ATP is absent, no filament movement is observed. Using this assay, one can show that the S1 fragment of myosin II is sufficient to bring about movement of actin filaments. This movement is caused by the tethered myosin S1 fragments (bound to the coverslip) trying to “move” toward the (+) end of a filament; thus the filaments move with the (−) end leading. The rate at which myosin moves an actin filament can be determined from video recordings of sliding-filament assays (Figure 17-23b).
[Part (b) courtesy James Spudich.]
EXPERIMENTAL FIGURE 17-23The sliding-filament assay is used to detect myosin-powered movement. (a) After myosin molecules are adsorbed onto the surface of a glass coverslip, excess unbound myosin is removed; the coverslip is then placed myosin-side-down on a glass slide to form a chamber through which solutions can flow. A solution of actin filaments, made visible and stable by staining with rhodamine-labeled phalloidin, is allowed to flow into the chamber. In the presence of ATP, the myosin heads “walk” toward the (+) ends of the actin filaments by the mechanism discussed later and illustrated in Figure 17-26. Because the myosin tails are immobilized, walking of the heads toward the (+) ends causes sliding of the filaments, which appear to be moving with their (−) ends leading the way. Movement of individual filaments can be observed in a fluorescence light microscope. (b) These photographs show the positions of three actin filaments (numbered 1, 2, 3) at 30-second intervals recorded by video microscopy. The rate of filament movement can be determined from such recordings.
[Part (b) courtesy James Spudich.]
All myosins have a domain related to the S1 domain of myosin II, comprising the head and neck domains, that is responsible for their motor activity. However, as we will see in a later section, the length of the neck domain and the numbers and types of light chains associated with it vary among myosin classes. The tail domain does not contribute to motility, but rather defines what is moved by the S1-related domain. Thus, as might be expected, the tail domains can be very different and are tailored to bind specific cargoes.