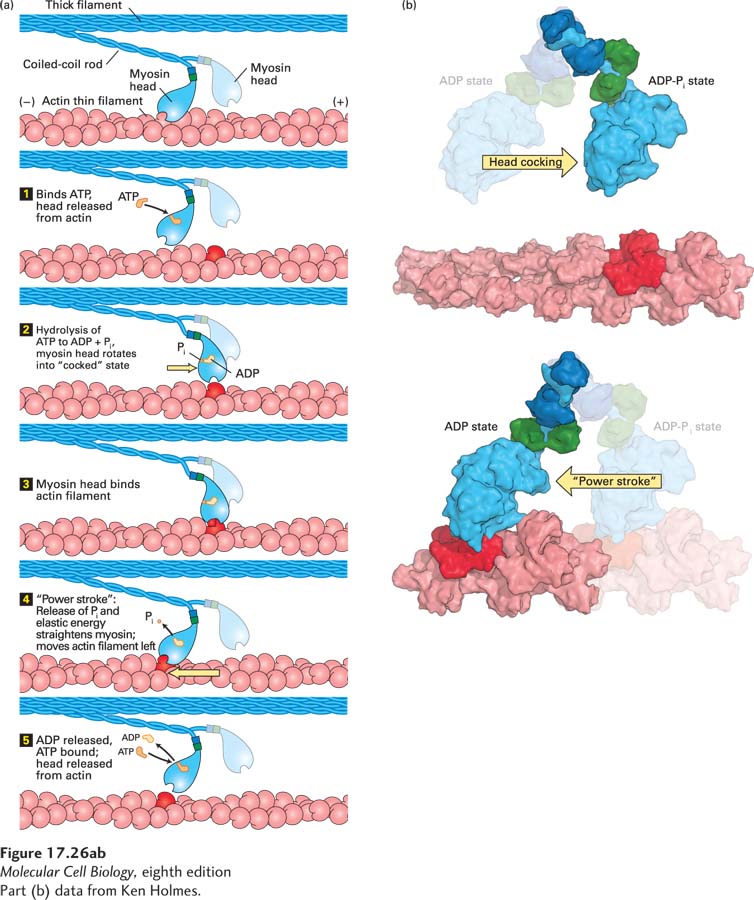
Conformational Changes in the Myosin Head Couple ATP Hydrolysis to Movement
Studies of muscle contraction provided the first evidence that myosin heads slide or walk along actin filaments. The unraveling of the mechanism of muscle contraction was greatly aided by the development of in vitro motility assays and single-molecule force measurements. On the basis of information obtained with these techniques and the three-dimensional structure of the myosin head (see Figure 17-22c), researchers developed a general model for how myosin harnesses the energy released by ATP hydrolysis to move along an actin filament (Figure 17-26). Because all myosins are thought to use the same basic mechanism to generate movement, we will ignore for the moment whether the myosin tail is bound to a vesicle or is part of a thick filament, as it is in muscle. The most important aspect of this model is that the hydrolysis of a single ATP molecule is coupled to each step taken by a myosin molecule along an actin filament.
[Part (b) data from Ken Holmes.]
FIGURE 17-26ATP-driven myosin movement along actin filaments. (a) In the absence of ATP, the myosin head is firmly attached to the actin filament. Although this state is very short-lived in living muscle, it is the state responsible for muscle stiffness in death (rigor mortis). Step 1: On binding ATP, the myosin head releases from the actin filament. Step 2: The head hydrolyzes the ATP to ADP and Pi, which induces a rotation in the head with respect to the neck. This “cocked state” stores the energy released by ATP hydrolysis as elastic energy, like a stretched spring. Step 3: Myosin in the “cocked” state is stable until it binds actin. Step 4: When it is bound to actin, the myosin head couples release of Pi with release of the elastic energy to move the actin filament. This movement is known as the “power stroke,” as it involves moving the actin filament with respect to the end of the myosin neck domain. Step 5: The head remains tightly bound to the actin filament until ADP is released and fresh ATP is bound by the head. See R. D. Vale and R. A. Milligan, 2002, Science288:88. (b) Molecular models of the conformational changes in the myosin head involved in “cocking” the head (upper panel) and after the power stroke (lower panel). The myosin light chains are shown in dark blue and green; the rest of the myosin head and neck are colored light blue, and actin is red. See S. Fischer et al., 2005, Proc. Natl. Acad. Sci. USA102:6873–6876.
[Part (b) data from Ken Holmes.]
How can myosin convert the chemical energy released by ATP hydrolysis into mechanical work? As mentioned above, the S1 head of myosin is an ATPase and thus has the ability to hydrolyze ATP into ADP and Pi. Biochemical analysis has revealed the mechanism of myosin movement (Figure 17-26a). In the absence of ATP, the head of myosin binds very tightly to F-actin. When ATP binds to the head, its affinity for F-actin is greatly reduced, and it releases from actin. The myosin head then hydrolyzes the ATP, and the hydrolysis products, ADP and Pi, remain bound to it. The energy provided by the hydrolysis of ATP induces a conformational change in the head that results in the head domain rotating with respect to the neck, assuming what is known as the “cocked” position (Figure 17-26b, top). In the absence of F-actin, release of Pi is exceptionally slow—the slowest part of the ATPase cycle. However, in the presence of actin, the head binds F-actin tightly, inducing both release of Pi and rotation of the head back to its original position, thus moving the actin filament relative to the neck domain (Figure 17-26b, bottom). In this way, binding to F-actin induces the movement of the head and the release of Pi, thereby coupling the two processes. This step is known as the power stroke. The head remains bound until the ADP leaves and a fresh ATP binds the head, releasing it from the actin filament. The cycle then repeats, and the myosin can move again against the filament.
How is the hydrolysis of ATP in the nucleotide-binding pocket of the head converted into force? The hydrolysis of ATP causes a small conformational change in the head domain. This small movement is amplified by a “converter” region at the base of the head, which acts like a fulcrum and causes the rodlike neck, also known as the lever arm, to rotate. This rotation is amplified by the neck domain, so the actin filament moves by a few nanometers (see Figure 17-26b).
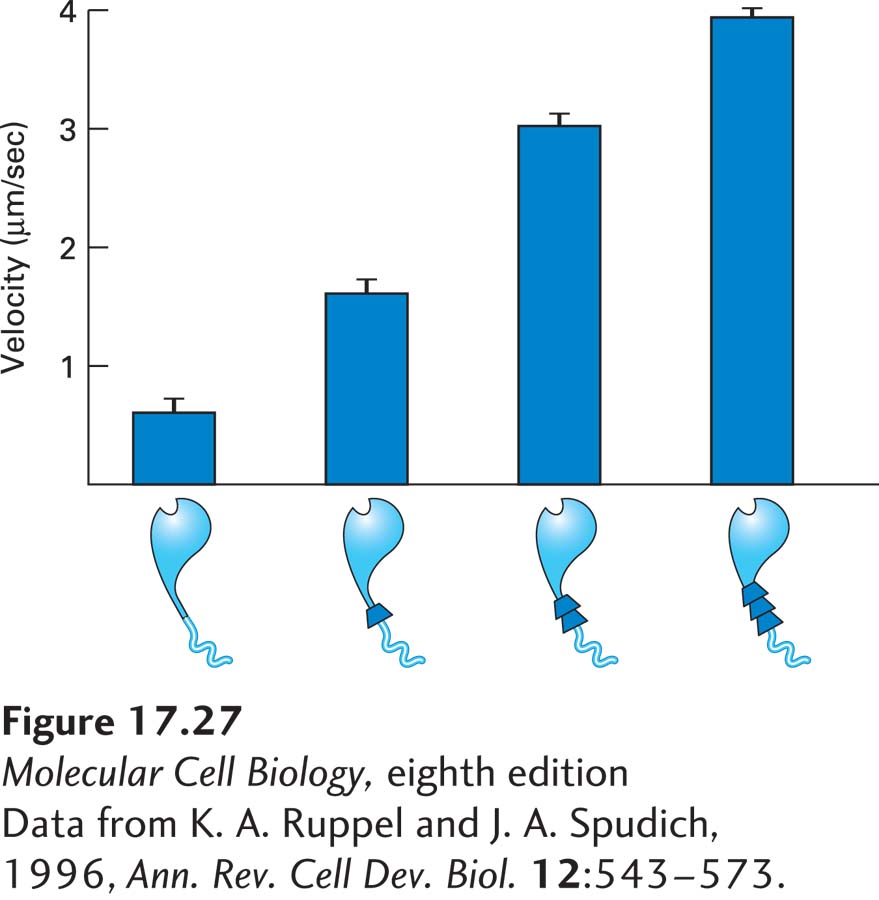
This model makes a strong prediction: the distance a myosin moves along actin during hydrolysis of one ATP—the myosin step size—should be proportional to the length of the neck domain. To test this prediction, mutant myosin molecules were constructed with neck domains of different lengths, and the rate at which they moved down an actin filament was determined. Remarkably, there was an excellent correlation between the length of the neck domain and the rate of movement (Figure 17-27).
Page 801
Page 802
[Data from K. A. Ruppel and J. A. Spudich, 1996, Ann. Rev. Cell Dev. Biol.12:543–573.]
EXPERIMENTAL FIGURE 17-27The length of the myosin II neck domain determines the rate of movement. To test the lever-arm model of myosin movement, investigators used recombinant DNA techniques to make myosin heads attached to neck domains of different lengths. The rate at which these myosins moved on actin filaments was determined. The longer the lever arm, the faster the myosin moved, supporting the proposed mechanism.
[Data from K. A. Ruppel and J. A. Spudich, 1996, Ann. Rev. Cell Dev. Biol.12:543–573.]