Myosin V–Bound Vesicles Are Carried Along Actin Filaments
In contrast to the contractile functions of myosin II filaments, the myosin V family of proteins, the most processive myosin motors known, transport cargo along actin filaments. In the next chapter we discuss how they can work together with microtubule motors to bring about transport of organelles. Although little is known about their functions in mammalian cells, myosin V motors are not unimportant: defects in a specific myosin V protein can cause severe diseases, such as seizures (see Figure 17-24).
Page 809
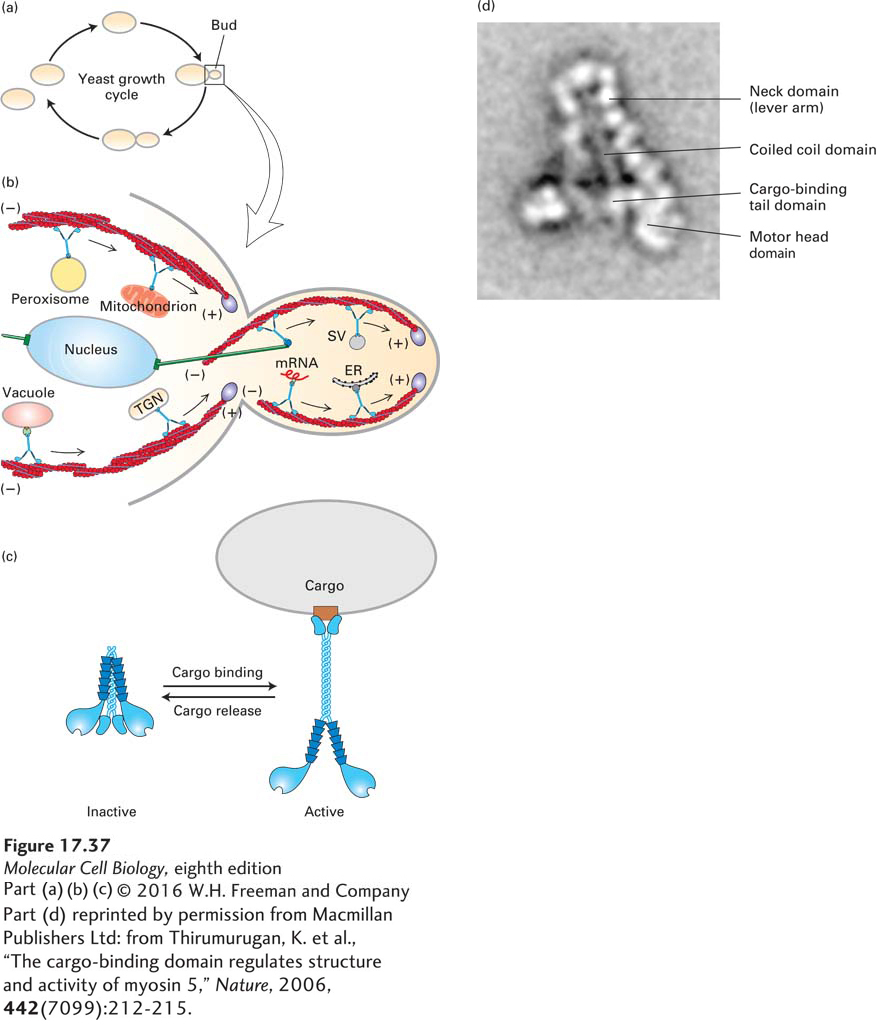
Much more is known about myosin V motors in more experimentally accessible and simpler systems such as the budding yeast. This well-studied organism grows by budding, which requires its secretory machinery to target newly synthesized materials to the growing bud (Figure 17-37a). Myosin V transports secretory vesicles along actin filaments at 3 µm/s into the bud. However, this is not the only function of myosin V proteins in yeast. At a later stage of the cell cycle, all the organelles have to be distributed between the mother and daughter cells. Remarkably, myosin V molecules in yeast provide the transport system for segregation of many organelles, including peroxisomes, mitochondria, vacuoles, endoplasmic reticulum, the trans-Golgi network, and even transport the ends of microtubules and some specific messenger RNAs into the bud (Figure 17-37b). Each of these organelles has a receptor to which the myosin V binds. The motor makes many delivery cycles, so it has to have a way to pick up, transport, and then deliver its organelle cargo. Recent work has shown that myosin V can exist in an inactive folded state in which the tail binds and inhibits the activity of the motor head domain, and an active state in which binding to cargo relieves the head-to-tail interaction (Figure 17-37c, d). How the motor releases its cargo upon delivery is poorly understood—in one case, the organelle receptor becomes degraded upon delivery of the organelle to its destination. Whereas budding yeast uses myosin V and polarized actin filaments in the transport of many organelles, animal cells, which are much larger, employ microtubules and their motors to transport many of the same organelles over the relatively longer distances involved. We discuss those transport mechanisms in the next chapter.
[Part (d) reprinted by permission from Macmillan Publishers Ltd: from Thirumurugan, K. et al., “The cargo-binding domain regulates structure and activity of myosin 5,” Nature, 2006, 442(7099):212-215.]
FIGURE 17-37Cargo movement by myosin V. (a) The yeast Saccharomyces cerevisiae (used in making bread, beer, and wine) grows by budding. Secretory vesicles are transported into the bud, which swells to about the size of the mother cell. Before cell division can occur, all the organelles must be segregated between the mother and the bud. The cell then undergoes cytokinesis to form two daughter cells. As shown in (b), all these transport processes depend on myosin V. This diagram of a medium-sized bud shows how myosin V transports secretory vesicles (SV) down actin cables nucleated by formins (purple) located at the bud tip and bud neck. Myosin V is also used to segregate organelles such as the vacuole (the yeast equivalent of a lysosome), peroxisomes, mitochondria, endoplasmic reticulum (ER), trans-Golgi network (TGN), and even selected mRNAs into the bud. Myosin V also binds the ends of cytoplasmic microtubules (green) to orient the nucleus in preparation for mitosis. See D. Pruyne et al., 2004, Ann. Rev. Cell Dev. Biol.20:559. (c) Myosin V is regulated by binding cargo. In the inactive folded state, the tails of myosin V bind and inactivate the motor head domains. Upon cargo binding, the head-to-tail interaction is alleviated, and the motor is now active to transport cargo. (d) Electron micrograph of inactive myosin V. Myosin V molecules were negatively stained and viewed in a transmission electron microscope. The image shown is the composite average from several individual images.
[Part (d) reprinted by permission from Macmillan Publishers Ltd: from Thirumurugan, K. et al., “The cargo-binding domain regulates structure and activity of myosin 5,” Nature, 2006, 442(7099):212-215.]
Page 810
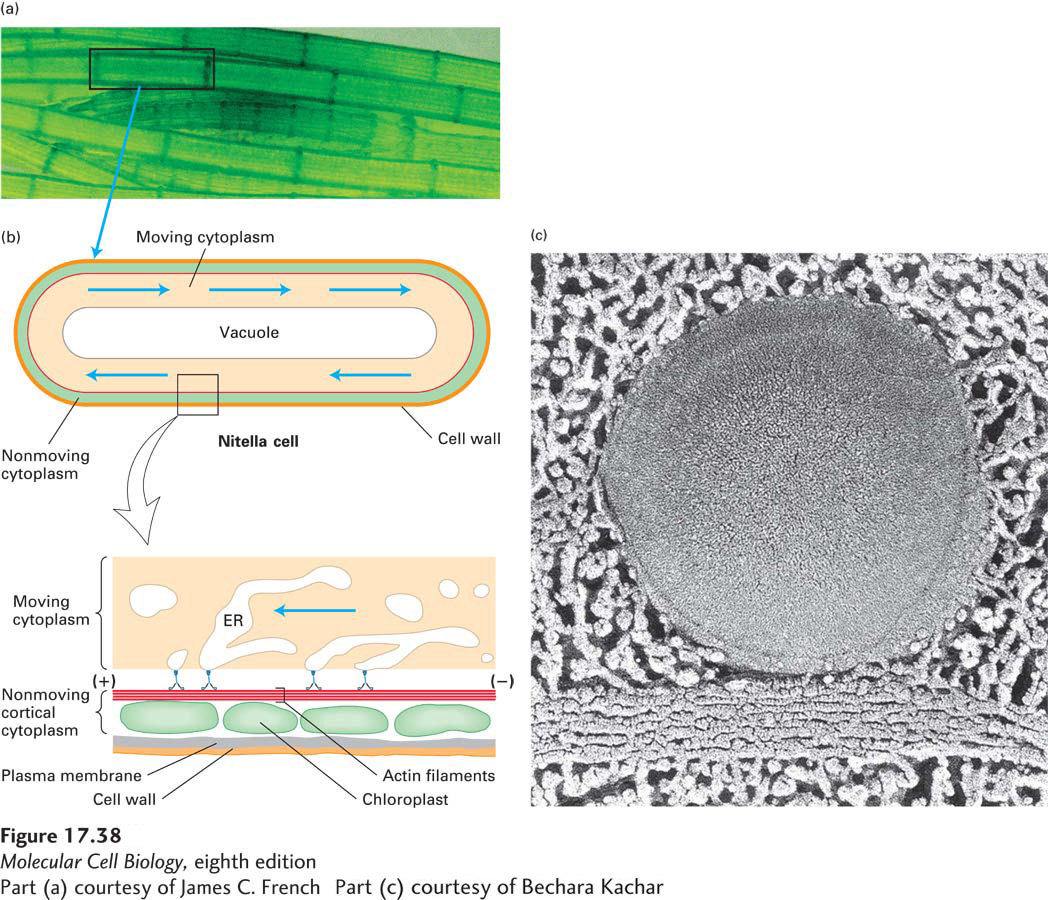
Perhaps the most dramatic use of myosin V is seen in giant green algae, such as Nitella and Chara. These algae can be readily found in ponds during the summer and their movement easily observed using a simple microscope. In their large cells, which can be as much as 2 cm in length, cytosol flows rapidly, at a rate approaching 4.5 mm per minute, in an endless loop around the inner circumference of the cell (Figure 17-38). This cytoplasmic streaming is a principal mechanism for distributing cellular metabolites, especially in large cells such as plant cells and amoebae. The algal cells have bundles of actin filaments aligned along the length of the cell, lying just above the stationary chloroplasts located adjacent to the membrane. The bulk cytosol is propelled by myosin V (also known as myosin XI in plants) attached to parts of the ER adjacent to the actin filaments. The flow rate of the cytosol in Nitella is about 15 times as fast as the movement produced by any other known myosin.
[Part (a) courtesy of James C. French; part (c) courtesy of Bechara Kachar.]
FIGURE 17-38Cytoplasmic streaming in cylindrical giant algae. (a) Cells of Nitella, a freshwater alga commonly found in ponds in the summer. The cytoplasmic movement, described below, can be readily observed with a simple microscope, so go find some Nitella (or related algae) and watch this amazing phenomenon! (b) The center of a Nitella cell is filled with a single large water-filled vacuole, which is surrounded by a layer of moving cytoplasm (blue arrows). A nonmoving layer of cortical cytoplasm filled with chloroplasts lies just under the plasma membrane (enlarged in bottom figure). On the inner side of this layer are bundles of stationary actin filaments (red), all oriented with the same polarity. A motor protein (blue), a plant myosin V, carries parts of the endoplasmic reticulum (ER) along the actin filaments. The movement of the ER network propels the entire viscous cytoplasm, including organelles that are enmeshed in the ER network. (c) Electron micrograph of the cortical cytoplasm showing a large vesicle connected to an underlying bundle of actin filaments.
[Part (a) courtesy of James C. French; part (c) courtesy of Bechara Kachar.]