G-Actin Monomers Assemble into Long, Helical F-Actin Polymers
Actin exists as a globular monomer called G-actin and as a filamentous polymer called F-actin, which is a linear chain of G-actin subunits. Each actin molecule contains a Mg2+ ion complexed with either ATP or ADP. In fact, actin is an ATPase, as it will hydrolyze ATP to ADP and Pi. The importance of the interconversion between the ATP and the ADP forms of actin is discussed below.
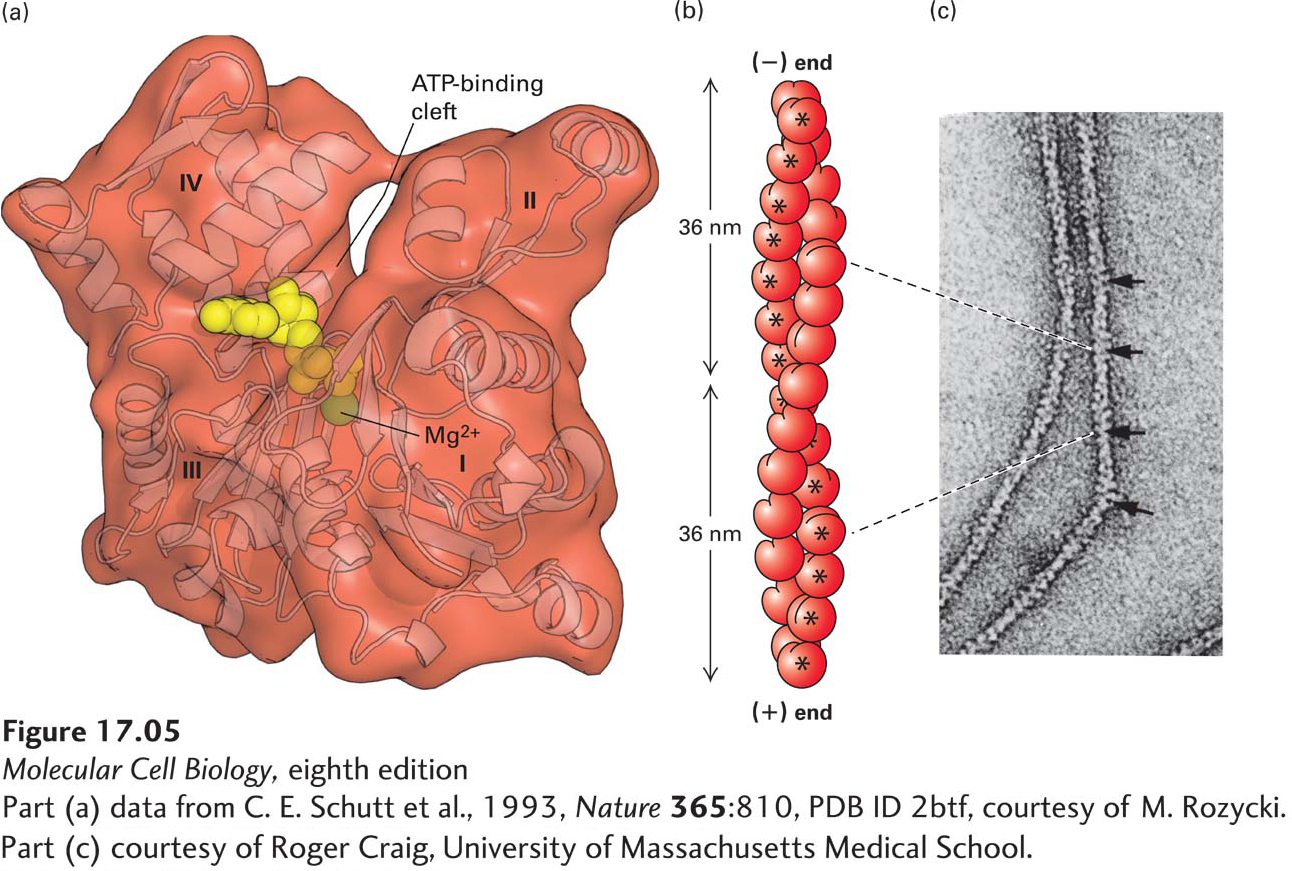
X-ray crystallographic analysis reveals that the G-actin monomer is separated into two lobes by a deep cleft (Figure 17-5a). At the base of the cleft is the ATPase fold, the site where ATP and Mg2+ are bound, which has structural similarity to the GTP-binding cleft of the GTPase molecular switches (see Figure 15-5). The floor of the cleft acts as a hinge that allows the lobes to flex relative to each other. When ATP or ADP is bound to G-actin, the nucleotide affects the conformation of the molecule; in fact, without a bound nucleotide, G-actin denatures very quickly. The addition of cations—Mg2+, K+, or Na+—to a solution of G-actin will induce the polymerization of G-actin into F-actin filaments. The process is reversible: F-actin depolymerizes into G-actin when the ionic strength of the solution is lowered. The F-actin filaments that form in vitro are indistinguishable from microfilaments seen in cells, indicating that F-actin is the major component of microfilaments.
[Part (a) data from C. E. Schutt et al., 1993, Nature365:810, PDB ID 2btf, courtesy of M. Rozycki. Part (c) courtesy of Roger Craig, University of Massachusetts Medical School.]
FIGURE 17-5Structures of monomeric G-actin and F-actin filaments. (a) Structure of the actin monomer (measuring 5.5 × 5.5 × 3.5 nm), which is divided by a central cleft into two approximately equal-sized lobes and four subdomains, numbered I–IV. ATP (red) binds at the bottom of the cleft and contacts both lobes (the yellow ball represents Mg2+). The N- and C-termini lie in subdomain I. (b) An actin filament appears as two strands of subunits. One repeating unit consists of 28 subunits (14 in each strand, indicated by * for one strand), covering a distance of 72 nm. The ATP-binding cleft of every actin subunit is oriented toward the same end of the filament. The end of a filament with an exposed binding cleft is the (−) end; the opposite end is the (+) end. (c) In the electron microscope, negatively stained actin filaments appear as long, flexible, and twisted strands of beaded subunits. Because of the twist, the filament appears alternately thinner (7-nm diameter) and thicker (9-nm diameter) (arrows). (The microfilaments visualized in a cell by electron microscopy are F-actin filaments plus any bound proteins.)
[Part (a) data from C. E. Schutt et al., 1993, Nature365:810, PDB ID 2btf, courtesy of M. Rozycki. Part (c) courtesy of Roger Craig, University of Massachusetts Medical School.]
Page 780
From the results of x-ray diffraction studies of actin filaments and from the actin monomer structure shown in Figure 17-5a, scientists have determined that the subunits in an actin filament are arranged in a helical structure (Figure 17-5b). In this arrangement, the filament can be considered as two helical strands wound around each other. Each subunit in the structure contacts one subunit above it and one below it in its own strand as well as two subunits in the other strand. The subunits in a single strand wind around the back of the other strand and repeat after 72 nm, or 14 actin subunits. Since there are two strands, the actin filament appears to repeat every 36 nm (see Figure 17-5b). When F-actin is viewed by electron microscopy after negative staining with uranyl acetate, it appears as a twisted string whose diameter varies between 7 and 9 nm (Figure 17-5c).