Other End-Binding Proteins Regulate Microtubule Disassembly
Mechanisms also exist for enhancing the disassembly of microtubules. Although most of the regulation of microtubule dynamics appears to happen at the (+) end, in some situations, such as in mitosis, it can occur at both ends.
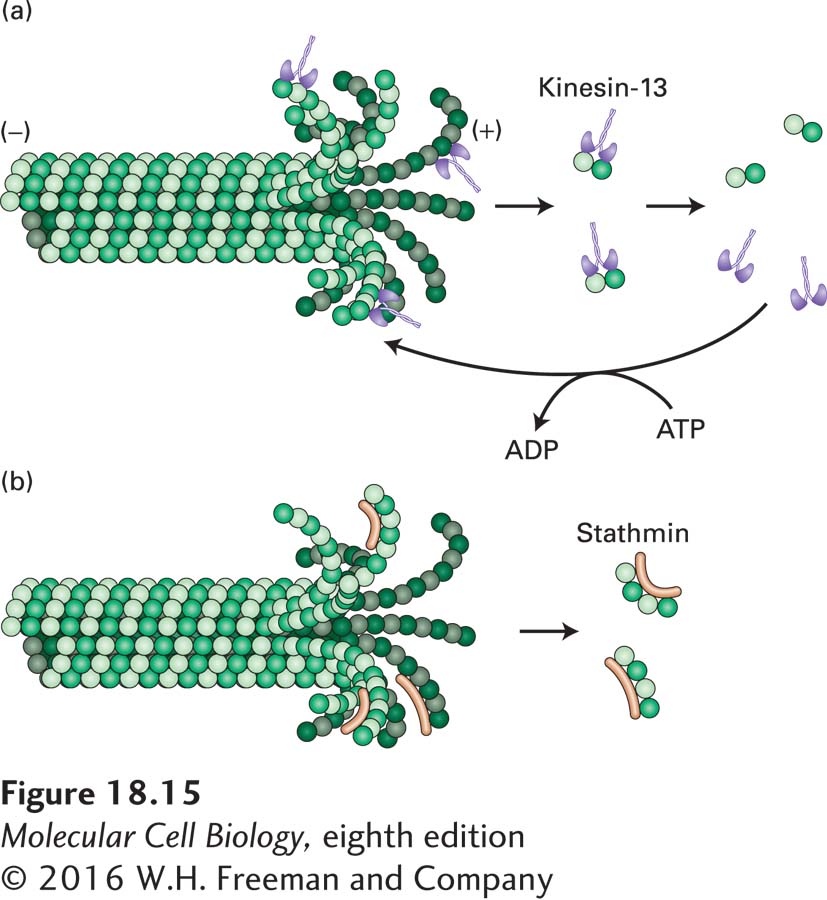
Various mechanisms for microtubule destabilization are known. One of these involves the kinesin-13 family of proteins. As we will see in Section 18.4, most kinesins are molecular motors, but the kinesin-13 proteins are a distinct class of kinesins that bind and curve the end of the tubulin protofilaments into the GDP-β-tubulin conformation. They then facilitate the removal of terminal tubulin dimers, thereby greatly enhancing the frequency of catastrophes (Figure 18-15a). They act catalytically in the sense that they need to hydrolyze ATP to remove sequential terminal tubulin dimers.
FIGURE 18-15Proteins that destabilize the ends of microtubules. (a) A member of the kinesin-13 family enriched at a microtubule end can enhance the disassembly of that end. [Although depolymerization of the (+) end is shown, kinesin-13 can also depolymerize the (−) end.] These proteins are ATPases, and ATP enhances their activity by dissociating them from the αβ-tubulin dimer. (b) Op18/stathmin binds selectively to curved protofilaments and enhances their dissociation from a microtubule end. This protein’s activity is inhibited by phosphorylation.
Another protein, known as Op18/stathmin, also enhances the rate of catastrophes. It was originally identified as a protein highly overexpressed in certain cancers; hence part of its name (Oncoprotein 18). Op18/stathmin is a small protein that binds two tubulin dimers in a curved, GDP-β-tubulin-like conformation (Figure 18-15b). It may function by enhancing the hydrolysis of the GTP in the terminal tubulin dimer and aiding in its dissociation from the end of the microtubule. As might be expected for a regulator of microtubule ends, it is subject to negative regulation by phosphorylation by a large variety of kinases. In fact, it has been found that Op18/stathmin is inactivated by phosphorylation near the leading edge of a motile cell, which contributes to preferential growth of microtubules toward the front of the cell.