Kinesin-1 Powers Anterograde Transport of Vesicles Down Axons Toward the (+) Ends of Microtubules
The protein responsible for anterograde organelle transport was first purified from axonal extracts. Researchers found that when they mixed three components—purified organelles from squid axons, an organelle-free cytosolic axonal extract, and taxol-stabilized microtubules—organelles could be seen moving on the microtubules in an ATP-dependent manner. However, if they omitted the axonal extract, the organelles neither bound nor moved along the microtubules. The researchers concluded that the extract contributes a protein that both attaches organelles to a microtubule and transports them along it—that is, a motor protein. Their strategy for purifying the motor protein was based on additional observations of organelles moving on microtubules. It was known that if ATP was hydrolyzed to ADP, the organelles fell off the microtubules. However, if the nonhydrolyzable ATP analog AMPPNP was added, the organelles remained associated with the microtubules, but did not move. These observations suggested that the motor protein linked the organelles to the microtubules very tightly in the presence of AMPPNP, but then was released from the microtubules when the AMPPNP was replaced by ATP, which was subsequently hydrolyzed to ADP. By looking for a protein that binds microtubules in the presence of AMPPNP and is released upon the addition of ATP, researchers were able to purify the motor protein, which they called kinesin.
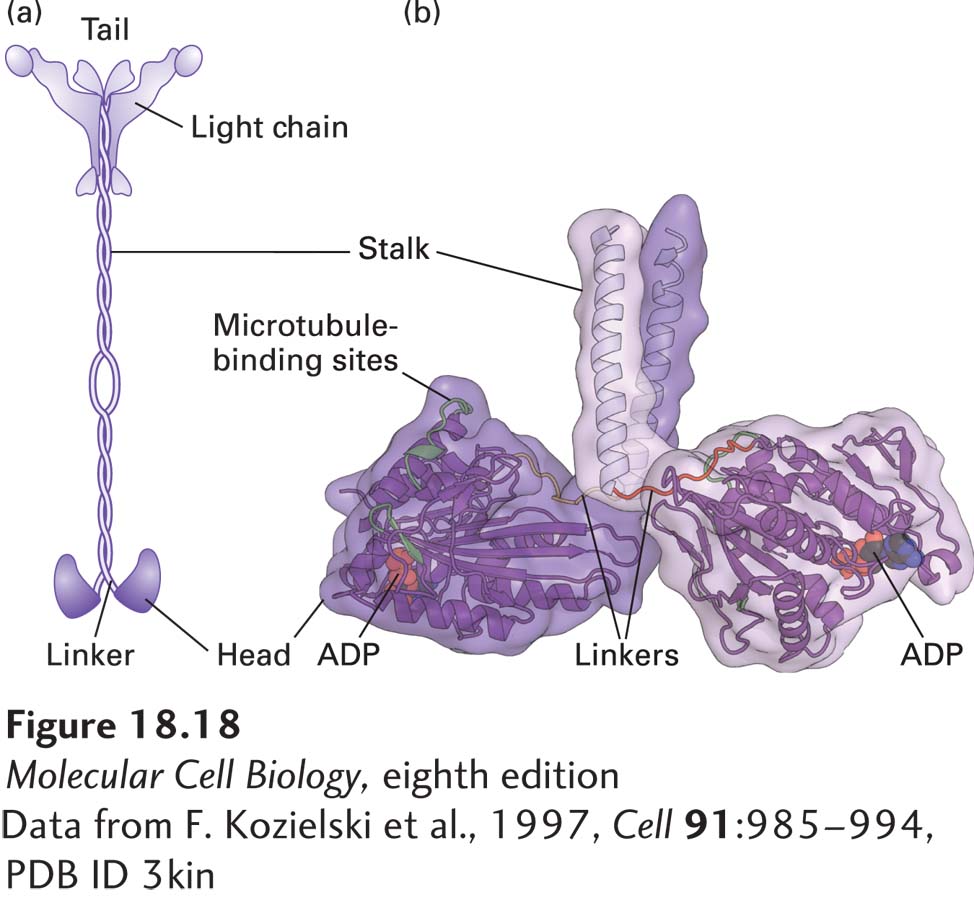
Kinesin-1 isolated from squid giant axons is a dimer of two heavy chains, each associated with a light chain, with a total molecular weight of about 380,000 Da. The molecule comprises a pair of globular N-terminal head domains connected by a short flexible linker domain to a long central stalk and terminating in a pair of small globular tail domains, which associate with the light chains (Figure 18-18). Each domain carries out a particular function: the head domain binds microtubules and ATP and is responsible for the motor activity of kinesin; the linker domain is critical for forward movement; the stalk domain is involved in dimerization through a coiled-coil interaction (see Figure 3-9) of the two heavy chains; and the tail domain is responsible for binding to receptors on the membranes of cargoes.
[Data from F. Kozielski et al., 1997, Cell91:985–994, PDB ID 3kin]
FIGURE 18-18Structure of kinesin-1. (a) Representation of kinesin-1 showing its two intertwined heavy chains, each with a motor domain in the head region. Each head is attached to the coiled-coil stalk by a flexible linker domain. Two light chains associate with the tail of the heavy chain. See R. D. Vale, 2003, Cell112:467. (b) X-ray structure of the kinesin heads with the microtubule-binding and nucleotide-binding sites (containing ADP) indicated, including the linkers and the beginning of the stalk region. See M. Thormahlen et al., 1998, J. Struct. Biol.122:30.
[Data from F. Kozielski et al., 1997, Cell91:985–994, PDB ID 3kin]
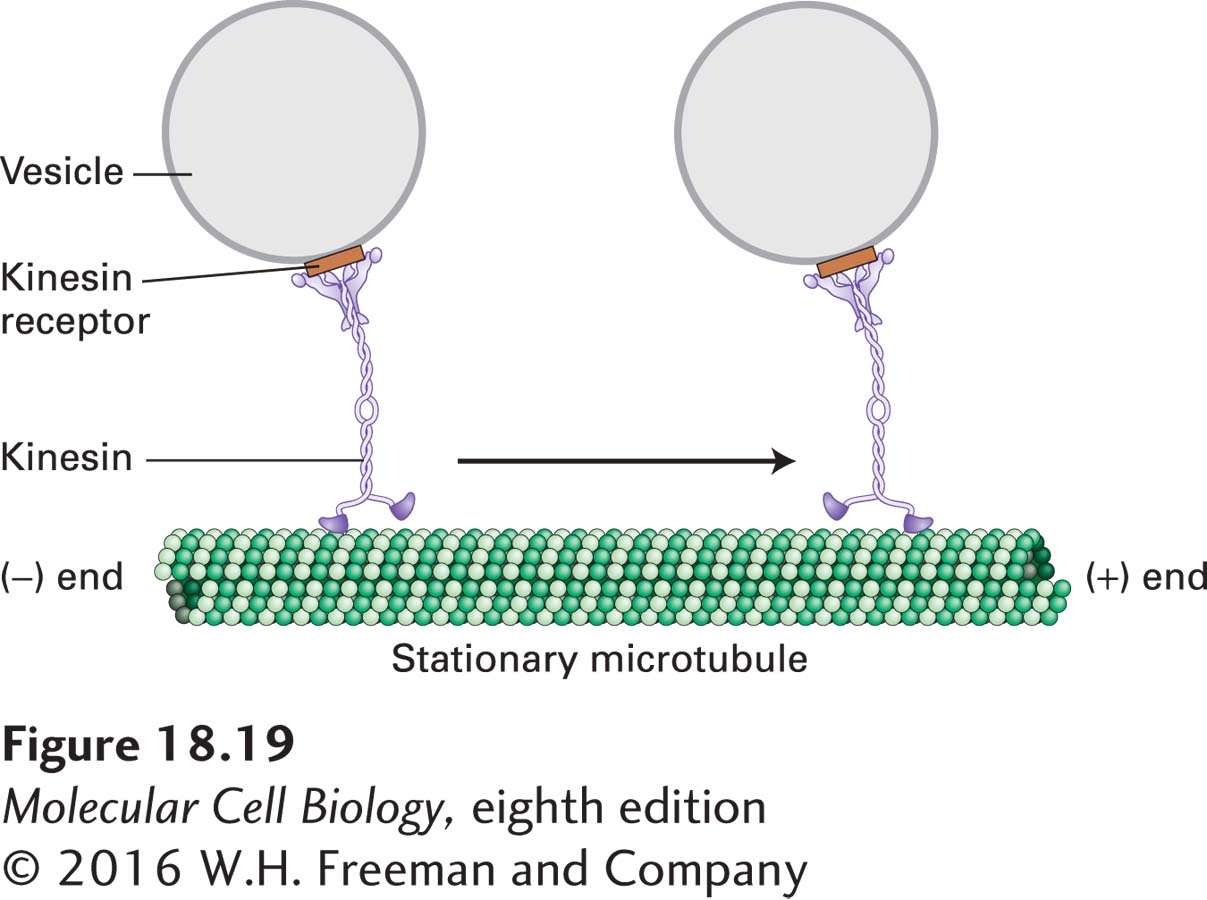
Kinesin-1-dependent movement of vesicles can be tracked by in vitro motility assays similar to those used to study myosin-dependent movements (see Figure 17-23). In one type of assay, a vesicle or a bead coated with kinesin-1 is added to a glass slide along with a preparation of stabilized microtubules and observed in a microscope. In the presence of ATP, the beads can be observed to move along a microtubule in one direction. Researchers found that beads coated with kinesin-1 always moved from the (−) to the (+) end of a microtubule (Figure 18-19). Thus kinesin-1 is a (+) end–directed microtubule motor protein, and additional evidence shows that it mediates anterograde transport of organelles along the axon.
FIGURE 18-19Model of kinesin-1-catalyzed vesicle transport. Kinesin-1 molecules, attached to receptors on the vesicle surface, transport the vesicles from the (−) end to the (+) end of a stationary microtubule. ATP is required for movement. See R. D. Vale et al., 1985, Cell40:559, and T. Schroer et al., 1988, J. Cell Biol. 107:1785.