How does kinesin-1 move down a microtubule? Optical trap and fluorescence-labeling techniques similar to those used to characterize myosin (see Figures 17-28 and 17-29) have been used to study how kinesin-1 moves down a microtubule and how ATP hydrolysis is converted into mechanical work. Such experiments demonstrate that it is a very processive motor, taking hundreds of “steps” walking “hand over hand” down a microtubule without dissociating from it. During this process, the double-headed molecule takes 8-nm steps from one tubulin dimer to the next, tracking down the same protofilament within the microtubule. This movement entails each individual head taking 16-nm steps. The two heads work in a highly coordinated manner so that one is always attached to the microtubule.
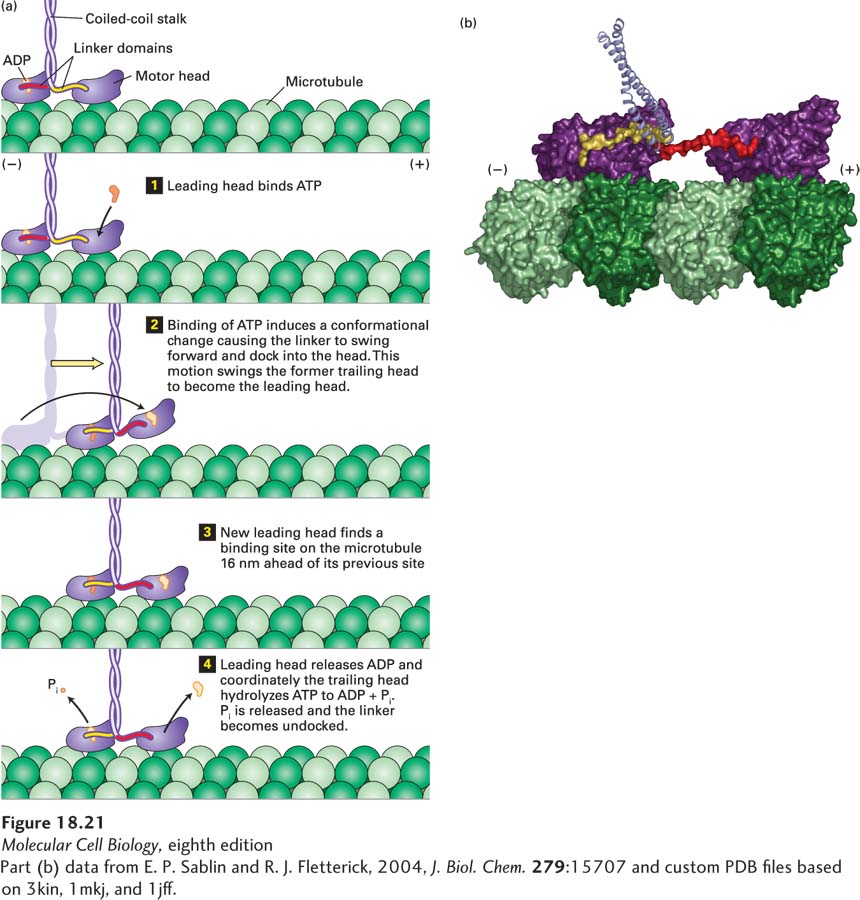
The ATP cycle of kinesin-1 movement is most easily understood if we begin just after the motor has taken a step (Figure 18-21a). At this point, the motor has a nucleotide-free leading head, which is strongly bound to a tubulin dimer in a protofilament, and an ADP-bound trailing head, which is weakly associated with the protofilament. ATP then binds to the leading head (Figure 18-21a, step 1), and this binding induces a conformational change in the linker domain so that instead of pointing backward, it swings forward and “docks” into its associated head. This swinging motion results in the linker domain rotating forward, and because it is attached to the trailing head, it swings the trailing head—like throwing a ballet dancer—into position to become the leading head (Figure 18-21a, step 2). The new leading head finds the next binding site on the microtubule (Figure 18-21a, step 3 and Figure 18-21b). The binding of the leading head to the microtubule induces the leading head to release ADP while the trailing head hydrolyzes ATP to ADP and Pi, releasing Pi (Figure 18-21a, step 4). ATP can now bind to the leading head to repeat the cycle and allow the protein to take another step down the microtubule.
[Part (b) data from E. P. Sablin and R. J. Fletterick, 2004, J. Biol. Chem.279:15707 and custom PDB files based on 3kin, 1mkj, and 1jff.]
FIGURE 18-21Kinesin-1 uses ATP to “walk” down a microtubule. (a) In this diagram, the two kinesin heads are shown with differently colored linker domains (yellow and red) to distinguish them. The cycle is shown starting after kinesin has taken a step, with the leading head tightly bound to the microtubule and not bound by any nucleotide, while the trailing head is weakly bound to the microtubule and has ADP bound. The leading head then binds ATP (step 1), which induces a conformational change that causes the yellow linker region to swing forward and dock into its associated head domain, thereby thrusting the trailing head forward (step 2). The new leading head now finds a binding site 16 nm down the microtubule, to which it binds weakly (step 3). The leading head now releases ADP and binds tightly to the microtubule, which induces the trailing head to hydrolyze ATP to ADP and Pi (step 4). Pi is released and the trailing head is converted into a weak binding state, and also releases the docked linker domain. The cycle now repeats itself for another step. See R. D. Vale and R. A. Milligan, 2000, Science288:88. (b) Structural model of two kinesin heads (purple) bound to a protofilament in a microtubule. The trailing head, at left, has bound ATP and has thrust the other head into the leading position. Notice how the linker domain (yellow) is docked into the trailing head, whereas the linker domain (red) of the leading head is still free.
[Part (b) data from E. P. Sablin and R. J. Fletterick, 2004, J. Biol. Chem.279:15707 and custom PDB files based on 3kin, 1mkj, and 1jff.]
Page 837
Two features of this cycle ensure that one head is always firmly bound to the microtubule. First, the head domain binds tightly to the microtubule in the nucleotide-free, ATP, and ADP + Pi states, but weakly in the ADP state. Second, the two heads communicate: when the leading head binds the microtubule and releases ADP, it is converted from a weak to a tight binding state. This change is communicated to the ATP-bound trailing head, which is tightly associated with the microtubule. The trailing head is stimulated to hydrolyze ATP, releasing Pi and converting to a weak binding state. Because this cycle requires that one head always be firmly attached to a tubulin dimer in a protofilament, kinesin-1 can take thousands of steps along a microtubule without disassociating from it.
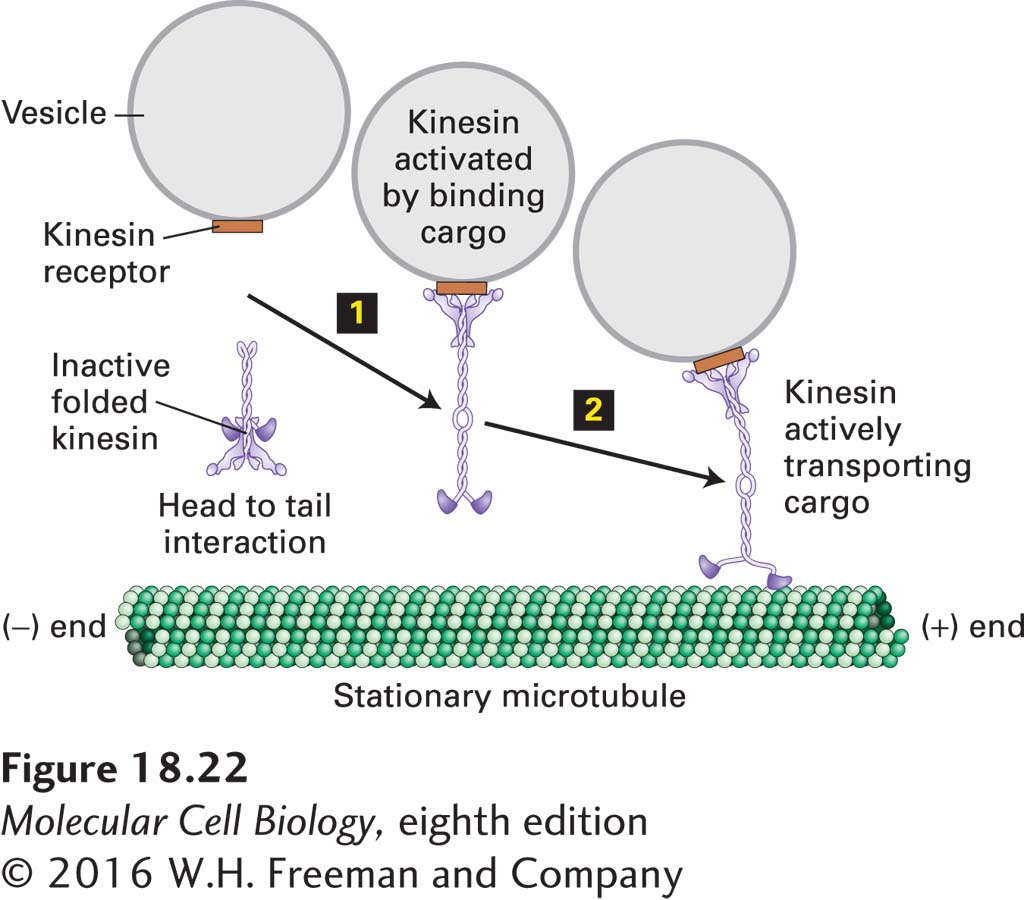
As a transporter of organelles, kinesin-1 must bind and transport the correct cargo. It does this through receptor proteins on the appropriate organelle that bind to the tail domain of the motor protein. Since kinesin-1 is an ATPase, it is important that this activity be inactivated to conserve energy when it is not needed, and also after releasing the organelle following transport. To accomplish this, kinesin-1 can fold into an inactive state in which the tail domain interacts with the motor head domain to inhibit both its ATPase activity and microtubule binding (Figure 18-22). When the tail binds the cargo-associated receptor protein, the motor unfolds and is activated (step 1). When the cargo-activated motor complex encounters a microtubule, it transports the organelle (step 2). Upon organelle delivery, kinesin-1 is released from the cargo and is inactivated by refolding into the head-to-tail inhibited state.
FIGURE 18-22Kinesin-1 is regulated by a head-to-tail interaction. In its inhibited form, the head of kinesin-1 folds back and interacts with the tail. This interaction inhibits the ATPase activity of kinesin-1. When the motor encounters an appropriate receptor, here shown on a vesicle, it unfolds (step 1) and is now able to transport the cargo toward the (+) end of a microtubule (step 2). It is not yet clear how the motor dissociates from the cargo when they arrive at the destination, but it involves cargo release and folding back into the inhibited state.
Page 838
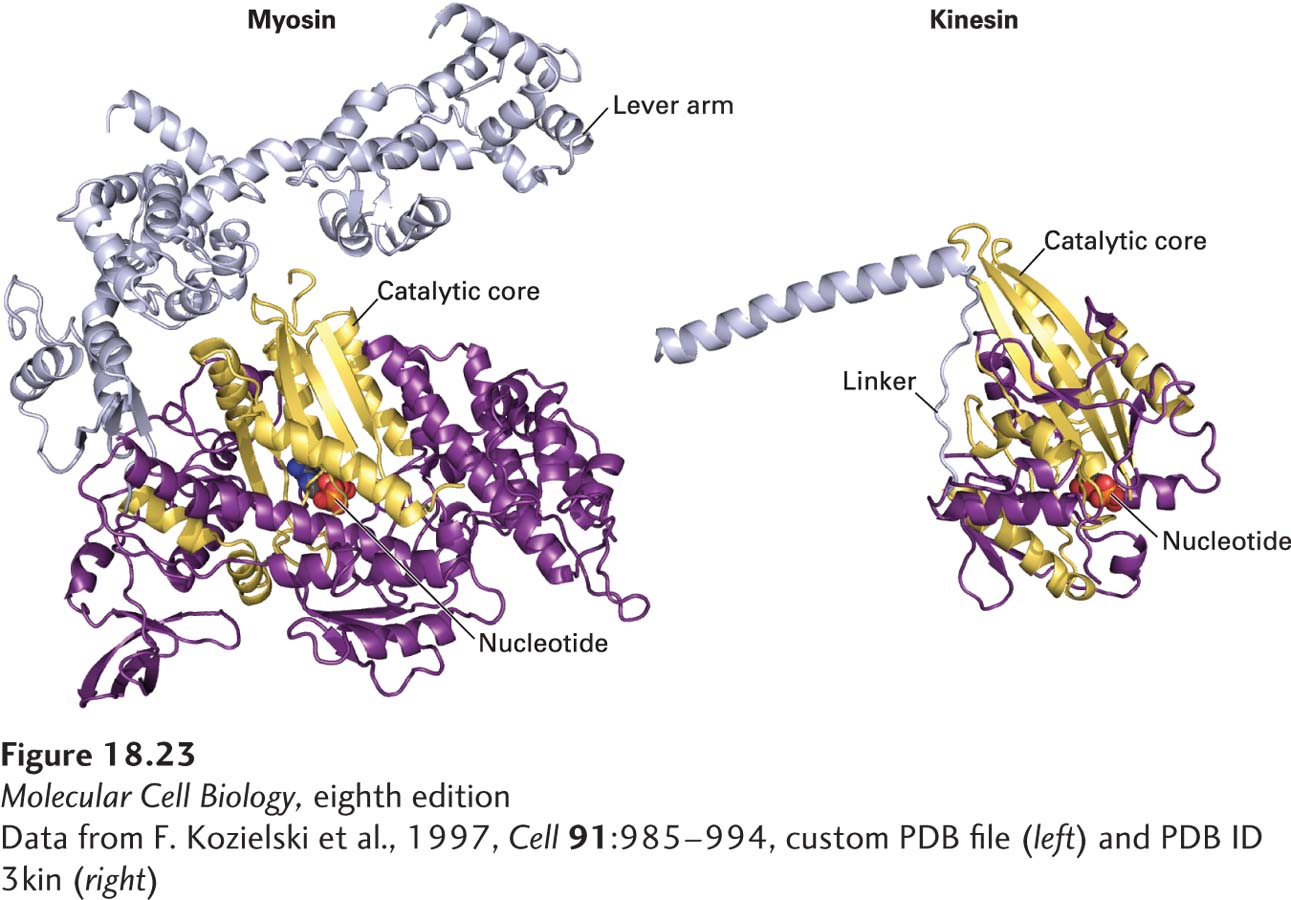
When the x-ray crystallographic structure of the kinesin head was determined, it revealed a major surprise—the catalytic core has the same overall structure as myosin’s (Figure 18-23)! This is the case despite the fact that there is no amino acid sequence conservation between the two proteins, arguing strongly that convergent evolution has twice generated a fold that can use the hydrolysis of ATP to generate work. Moreover, the same type of three-dimensional structure is seen in small GTP-binding proteins, such as Ras, that undergo a conformational change upon GTP hydrolysis (see Figure 15-7).
[Data from F. Kozielski et al., 1997, Cell91:985-994, custom PDB file (left) and PDB ID 3kin (right)]
FIGURE 18-23Convergent structural evolution of the ATP-binding cores of myosin and kinesin heads. The common catalytic cores of myosin and kinesin are shown in yellow, the nucleotide in red, and the lever arm (for myosin-II) and linker domain (for kinesin-1) in light purple. See R. D. Vale and R. A. Milligan, 2000, Science288:88.
[Data from F. Kozielski et al., 1997, Cell91:985-994, custom PDB file (left) and PDB ID 3kin (right)]