Kinesins and Dyneins Cooperate in the Transport of Organelles Throughout the Cell
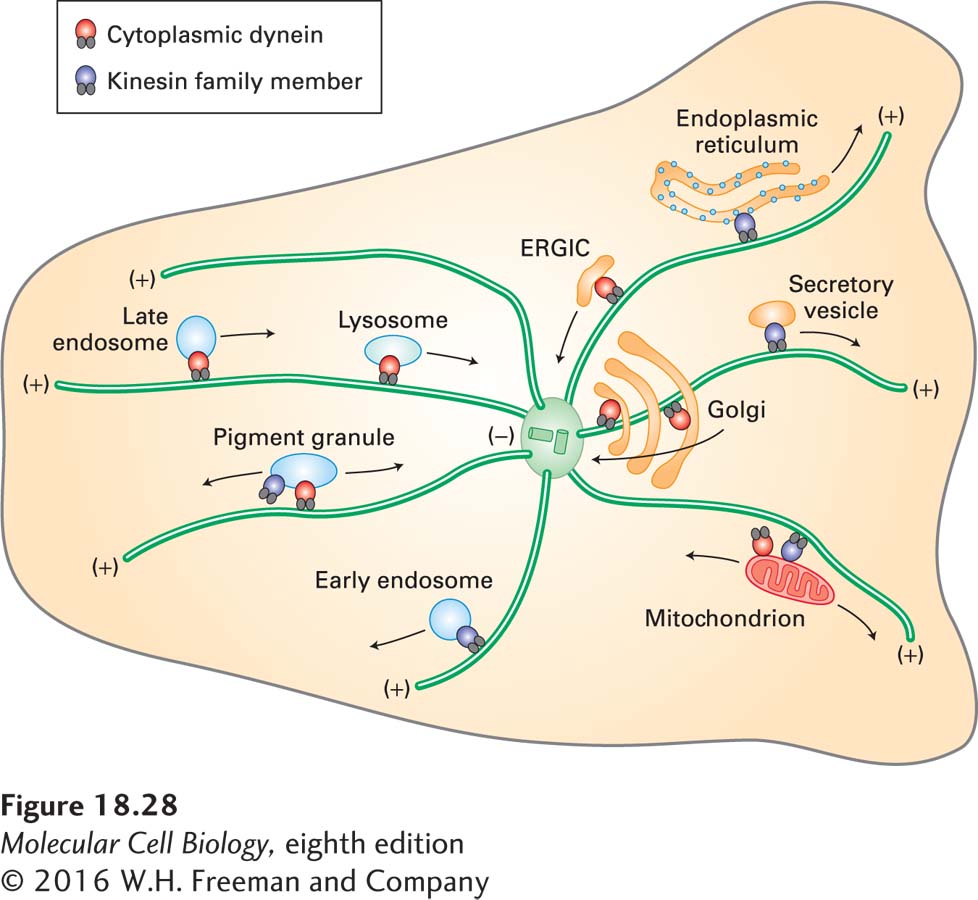
Both dynein and kinesin family members play important roles in the microtubule-dependent organization of organelles in cells (Figure 18-28). Because the orientation of microtubules is fixed by the MTOC, the direction of transport—toward or away from the cell center—depends on the motor protein. For example, the Golgi complex collects in the vicinity of the centrosome, where the (−) ends of microtubules lie, and is driven there by dynein-dynactin. In addition, secretory cargo emerging from the endoplasmic reticulum is transported to the Golgi by dynein-dynactin. Conversely, the endoplasmic reticulum is spread throughout the cytoplasm and is transported there by kinesin-1, which moves toward the peripheral (+) ends of microtubules. Some organelles of the endocytic pathway are associated with dynein-dynactin, including late endosomes and lysosomes. Kinesins have been shown to transport mitochondria as well as nonmembranous cargoes such as specific mRNAs encoding proteins that need to be localized during development.
FIGURE 18-28Organelle transport by microtubule motors. Cytoplasmic dyneins (red) mediate retrograde transport of organelles toward the (−) ends of microtubules (cell center); kinesins (purple) mediate anterograde transport toward the (+) ends (cell periphery). Most organelles have one or more microtubule-based motors associated with them. It should be noted that the association of motors with organelles varies by cell type, so some of these associations may not exist in all cells, whereas others not shown here also exist. ERGIC = ER-to-Golgi intermediate compartment.
We have seen how kinesin-1 transports organelles in an anterograde direction down axons. What happens to the motor when it gets to the end of the axon? The answer is that it is carried back in a retrograde direction on organelles transported by cytoplasmic dynein. Thus kinesin-1 and dynein can associate with the same organelle, and a mechanism must exist that turns one motor off while activating the other, although such mechanisms are not yet fully understood.
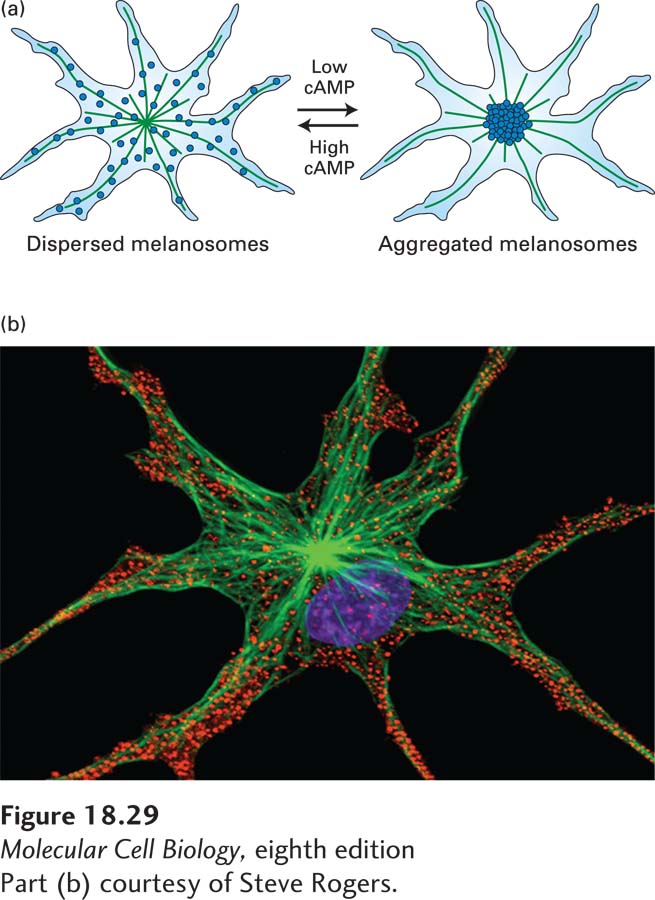
Much of what we know about the regulation of microtubule-based organelle transport comes from studies using fish (e.g., angelfish) or frog melanophores. Melanophores are cells of the vertebrate skin that contain hundreds of dark melanin-filled pigment granules called melanosomes. Melanophores either have their melanosomes dispersed, in which case they make the skin darker, or aggregated at the cell center, which makes the skin paler (Figure 18-29). These changes in skin color, mediated by neurotransmitters in the fish and regulated by hormones in the frog, serve to camouflage the fish and enhance social interactions in the frog. The movement of the melanosomes is mediated by changes in intracellular cAMP and is dependent on microtubules. Studies investigating which motors are involved have shown that melanosome dispersion requires kinesin-2, whereas melanosome aggregation requires cytoplasmic dynein-dynactin. The first hints of how these activities might be coordinated came from the finding that overexpression of dynamitin inhibited melanosome transport in both directions. This surprising result was explained when it was found that dynactin binds not only to cytoplasmic dynein, but also to kinesin-2—and may coordinate the activity of the two motors.
[Part (b) courtesy of Steve Rogers.]
FIGURE 18-29Movement of pigment granules in frog melanophores. (a) Diagram of the microtubule-based reorganization of melanosomes according to the level of cAMP. Melanosomes are aggregated by cytoplasmic dynein and dispersed by kinesin-2. (b) Melanosomes (red) in the dispersed state as seen by immunofluorescence microscopy. Microtubules are labeled green, and the DNA in the nucleus is labeled blue.
[Part (b) courtesy of Steve Rogers.]
Page 842
The association of dynein and kinesin-2 with the same organelle is not limited to melanosomes; it has recently been suggested that these motors may cooperate to appropriately localize late endosomes, lysosomes, and mitochondria in some cells. Thus the association of organelles with a number of distinct motors is not the exception, but an emerging theme.