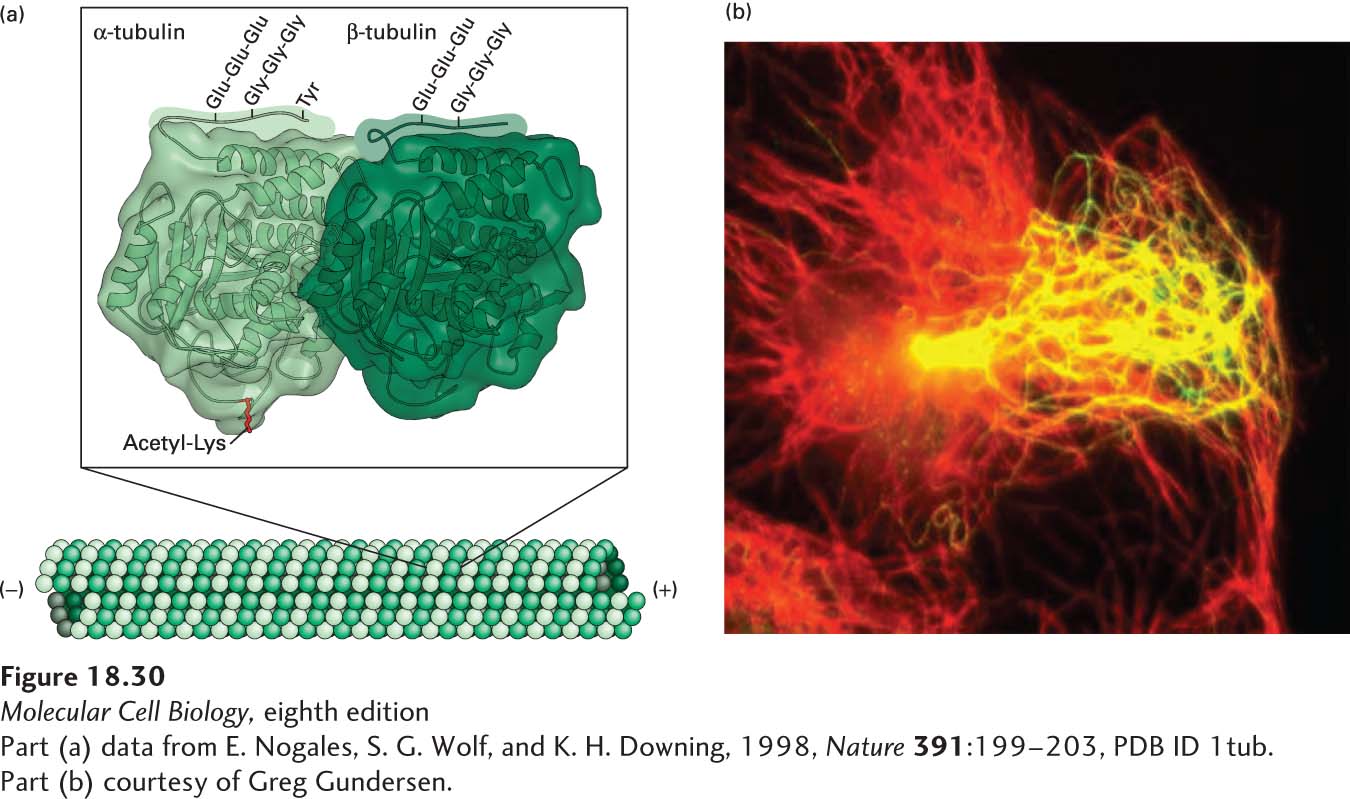
Tubulin Modifications Distinguish Different Classes of Microtubules and Their Accessibility to Motors
The stability and functions of different classes of microtubules are influenced by post-translational modifications. Although multiple types of modifications have been detected, we restrict our discussion here to those that are the best understood—lysine acetylation, detyrosylation, polyglutamylation, and polyglycylation (Figure 18-30a)—and their functional consequences.
[Part (a) data from E. Nogales, S. G. Wolf, and K. H. Downing, 1998, Nature391:199–203, PDB ID 1tub. Part (b) courtesy of Greg Gundersen.]
EXPERIMENTAL FIGURE 18-30Post-translational modifications of tubulin affect the stability and function of microtubules. (a) Structure of α- and β-tubulin, showing the sites of lysine acetylation on the inner surface of the microtubule and polyglutamylation, polyglycylation, and detyrosylation on the outer surface. Note that polyglutamylation and polyglycylation are believed to be mutually exclusive, so they would not normally occur at the same time. See J. W. Hamond, D. Cai, and K. J. Verhey, 2008, Curr. Opin. Cell Biol.20:71–76. (b) Detyrosylated microtubules are preferentially oriented toward the leading edge of a moving cell. A cell migrating toward the right is stained for total microtubules (red) and detyrosylated microtubules (green). The resulting merged image shows the detyrosylated microtubules enriched toward the front of the cell in yellow, which is a combination of red and green.
[Part (a) data from E. Nogales, S. G. Wolf, and K. H. Downing, 1998, Nature391:199–203, PDB ID 1tub. Part (b) courtesy of Greg Gundersen.]
Page 843
Two of these modifications are found only on α-tubulin and are absent from β-tubulin. The first is the acetylation of the ε-amino group of a specific lysine residue of α-tubulin that lies on the inside of the microtubule; microtubules with this acetylated lysine are found in stable microtubule structures such as centrioles, basal bodies, and primary cilia (primary cilia are discussed in Section 18.5). Indeed, cells unable to acetylate tubulin have defective primary cilia, whereas cells in which the acetylation cannot be removed have unusually stable primary cilia. The second modification of α-tubulin relates to its C-terminal tyrosine. This tyrosine can be specifically removed by a carboxypeptidase that functions only when bound to the surface of a microtubule, where it sequentially removes the C-terminal tyrosines from α-tubulin subunits. Such detyrosylated microtubules are more stable, as they are more resistant to depolymerization by the kinesin-13 family of depolymerizers. Moreover, in migrating cells, these more stable microtubules are generally oriented toward the front of the cell. When a detyrosylated microtubule depolymerizes, the α-tubulin subunit of the αβ-dimer has the C-terminal tyrosine added back by a tyrosine ligase that acts only on soluble tubulin, and the αβ-tubulin dimer can now be used during elongation of another growing microtubule.
The C-terminal regions of both α- and β-tubulin are very rich in glutamic acid residues, and specific enzymes can modify these residues. These modifications occur only after assembly of the microtubule. The tubulin tails can be modified by polyglutamylation, in which a chain of glutamic acid residues is linked to a specific glutamate residue, or by polyglycylation, in which a chain of glycine residues is added to a different glutamate residue. At present it is believed that these two modifications are mutually exclusive, so that if a tubulin subunit is modified by polyglycylation, it is protected from polyglutamylation, and vice versa. Like detyrosylation, polyglutamylation can enhance microtubule stability.
These post-translational modifications of tubulin not only affect microtubule stability, but can also influence the ability of molecular motors to interact with microtubules (Figure 18-30b). Kinesin-1 associates preferentially with detyrosylated and acetylated microtubules, so these modifications may be important in recruiting this motor for axonal transport in neurons. As we mentioned in Figure 18-5e, neurons have different microtubule organizations in their dendrites and their axons. The microtubules in the axon are stabilized by acetylation and detyrosylation, which allows kinesin-1 to associate preferentially with them for axonal transport. Polyglutamylation has a key role in the beating of cilia and flagella, which we discuss in the next section.
Page 844
The research elucidating the effects of post-translational modifications of tubulin on microtubule function and microtubule-based motors is all quite recent; we can expect future studies to reveal multiple “codes” that distinguish different classes of microtubules and specialize them for specific functions.