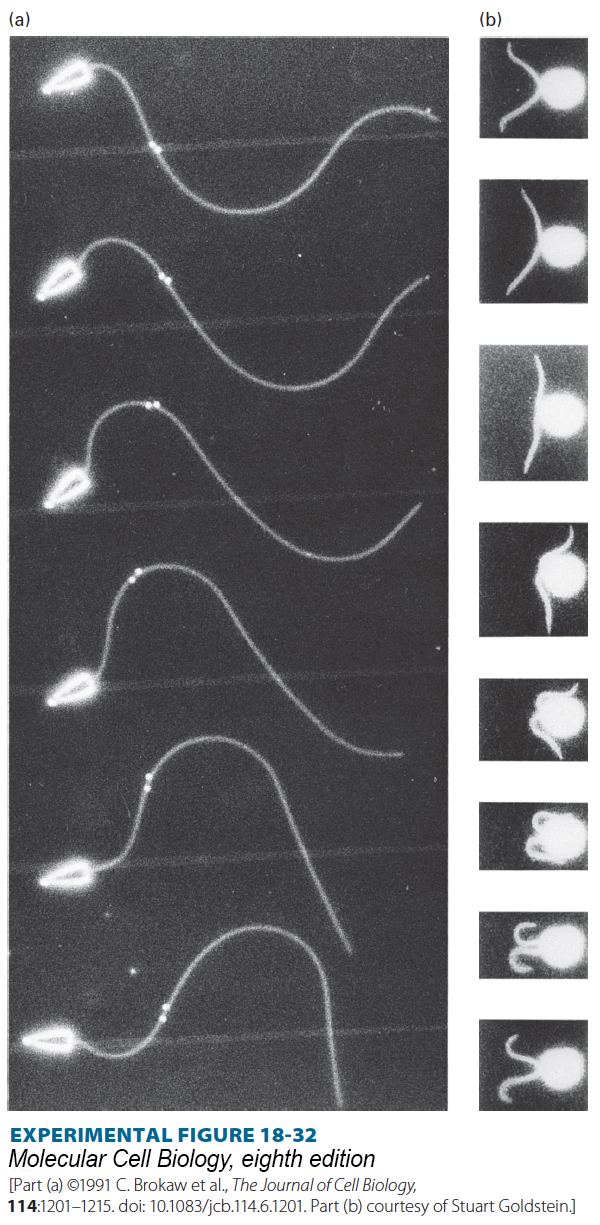
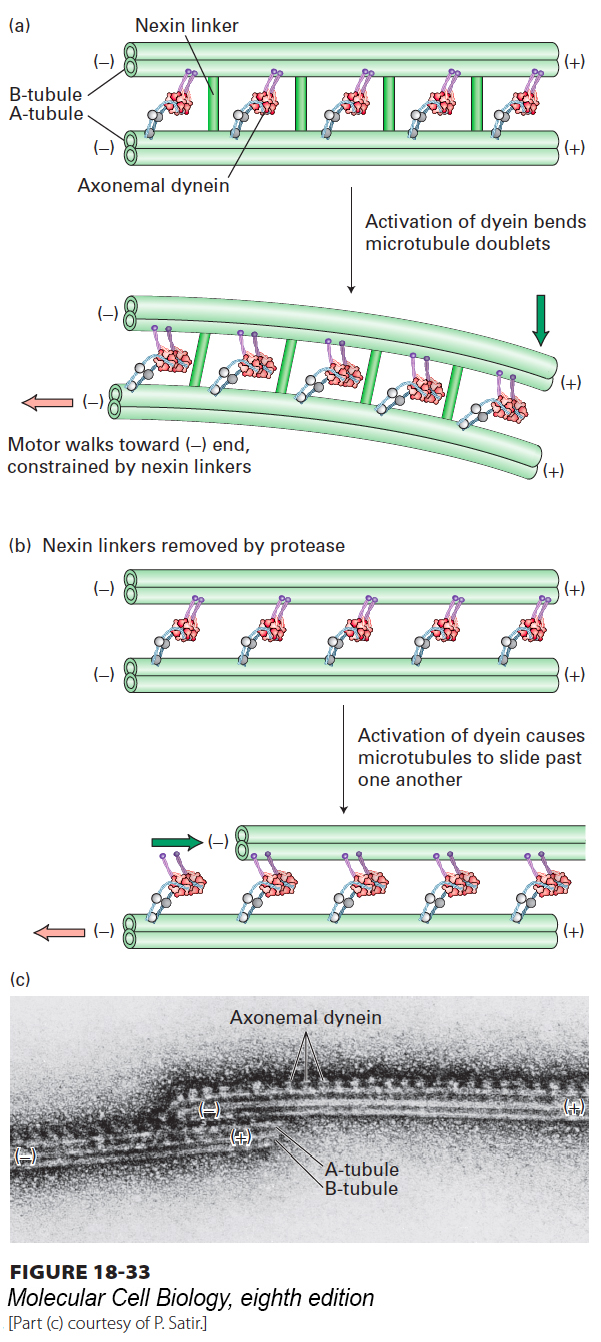
Ciliary and Flagellar Beating Are Produced by Controlled Sliding of Outer Doublet Microtubules
Cilia and flagella are motile structures because activation of the axonemal dynein motors causes them to bend. A close examination of their movement using video microscopy reveals that a bend starts at the base of a cilium or flagellum and then propagates along the structure (Figure 18-32). A clue to how this occurs came from studies of isolated axonemes. In these classic experiments, axonemes were gently treated with a protease that cleaves only the nexin linkers. When ATP was added to the treated axonemes, the doublet microtubules slid past one another as dynein, attached to the A tubule of one doublet, “walked” down the B tubule of the adjacent doublet (Figure 18-33b, c). In an axoneme with intact nexin linkers, the action of dynein induces flagellar bending because the microtubule doublets are connected to one another (Figure 18-33a).
EXPERIMENTAL FIGURE 18-32Video microscopy shows flagellar movements that propel sperm and Chlamydomonas forward. In both cases, the cells are moving to the left. (a) In the typical sperm flagellum, successive waves of bending originate at the base and are propagated out toward the tip; these waves push against the surrounding fluid and propel the cell forward. Captured in this multiple-exposure sequence, a bend at the base of the flagellum in the first (top) frame has moved distally halfway along the flagellum by the last frame. A pair of gold beads attached to the flagellum are seen to slide apart as the bend moves through their region. (b) Beating of the two flagella on Chlamydomonas occurs in two stages, called the effective stroke (top three frames) and the recovery stroke (remaining frames). The effective stroke pulls the organism through the water. During the recovery stroke, a different wave of bending moves outward from the bases of the flagella, pushing the flagella along the surface of the cell until they reach the position to initiate another effective stroke. Beating commonly occurs 5–10 times per second.
FIGURE 18-33Ciliary and flagellar bending is mediated by axonemal dynein. (a) Axonemal dynein attached to an A tubule of an outer doublet pulls on the B tubule of the adjacent doublet trying to move toward the (−) end. Because the adjacent tubules are tethered by nexin, the force generated by dynein bends the cilium or flagellum. (b) Experimental evidence for the model in (a). When the nexin linkers are cleaved with a protease and ATP added to induce dynein activity, the microtubule doublets slide past one another. (c) Electron micrograph of two doublet microtubules in a protease-treated axoneme incubated with ATP. In the absence of cross-linking proteins, doublet microtubules slide excessively. The dynein arms can be seen projecting from A tubules and interacting with B tubules of the left microtubule doublet.
[Part (c) courtesy of P. Satir.]
Page 845
How specific subsets of dynein are activated and how a wave of activation is propagated down the axoneme are not yet fully understood, but post-translational modifications of tubulin may play a role. Recall from Section 18.4 that post-translational modifications of tubulin subunits can affect the interactions between microtubules and motor proteins. The B tubules of the outer axoneme doublets are often polyglutamylated, and this modification strongly affects the interaction of the inner-arm dynein with the B tubule. Because inner-arm dynein motors mainly affect the waveform of the ciliary beat, it is this aspect of ciliary function that is compromised in mutants unable to undergo polyglutamylation.