Intraflagellar Transport Moves Material Up and Down Cilia and Flagella
Although axonemal dynein is involved in bending cilia and flagella, another type of motility has been observed in these structures more recently. Careful examination of flagella on the biflagellate green alga Chlamydomonas reinhardtii revealed cytoplasmic particles moving at a constant speed of about 2.5 µm/s toward the tip of a flagellum (anterograde movement) and other particles moving at about 4 µm/s from the tip to the base (retrograde movement). This movement, known as intraflagellar transport (IFT), occurs in both cilia and flagella. Light and electron microscopy revealed that the particles move between the outer doublet microtubules and the plasma membrane (Figure 18-34). Analysis of algal mutants demonstrated that the anterograde movement is powered by kinesin-
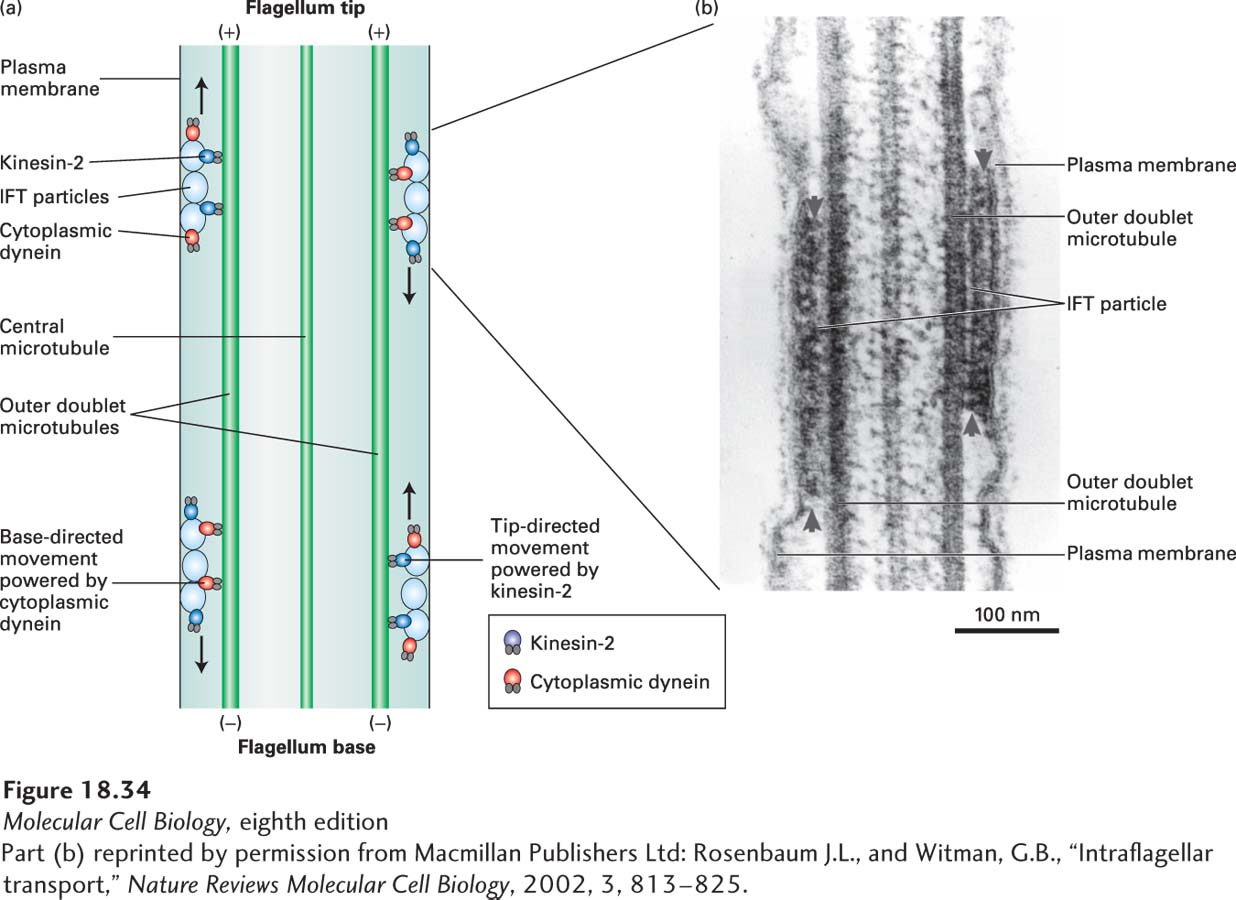
Page 846
Page 847
The anterograde and retrograde IFT particles transported in Chlamydomonas flagella have been isolated and their composition determined. They consist of two distinct protein complexes, called IFT complex A and IFT complex B. By analyzing the phenotypes of cells having mutations affecting these complexes, it has been found that complex B is necessary for anterograde IFT, whereas complex A is important for retrograde IFT. Despite this segregation of function, both complexes are transported in both directions. All the components of IFT particles have homologs in organisms containing cilia, such as nematodes, fruit flies, mice, and humans, but these particles are absent from the genomes of yeasts and plants that lack cilia, suggesting that they are specific to IFT.
What is the function of IFT? Because all the microtubules in a flagellum have their growing (+) ends at the tip, that is the site where new tubulin subunits and flagellar structural proteins are added. Moreover, even in cells with flagella of uniform length, the microtubules are turning over, with assembly and disassembly occurring at the flagellar tip. In cells defective for kinesin-