Microtubules Are Assembled from MTOCs to Generate Diverse Configurations
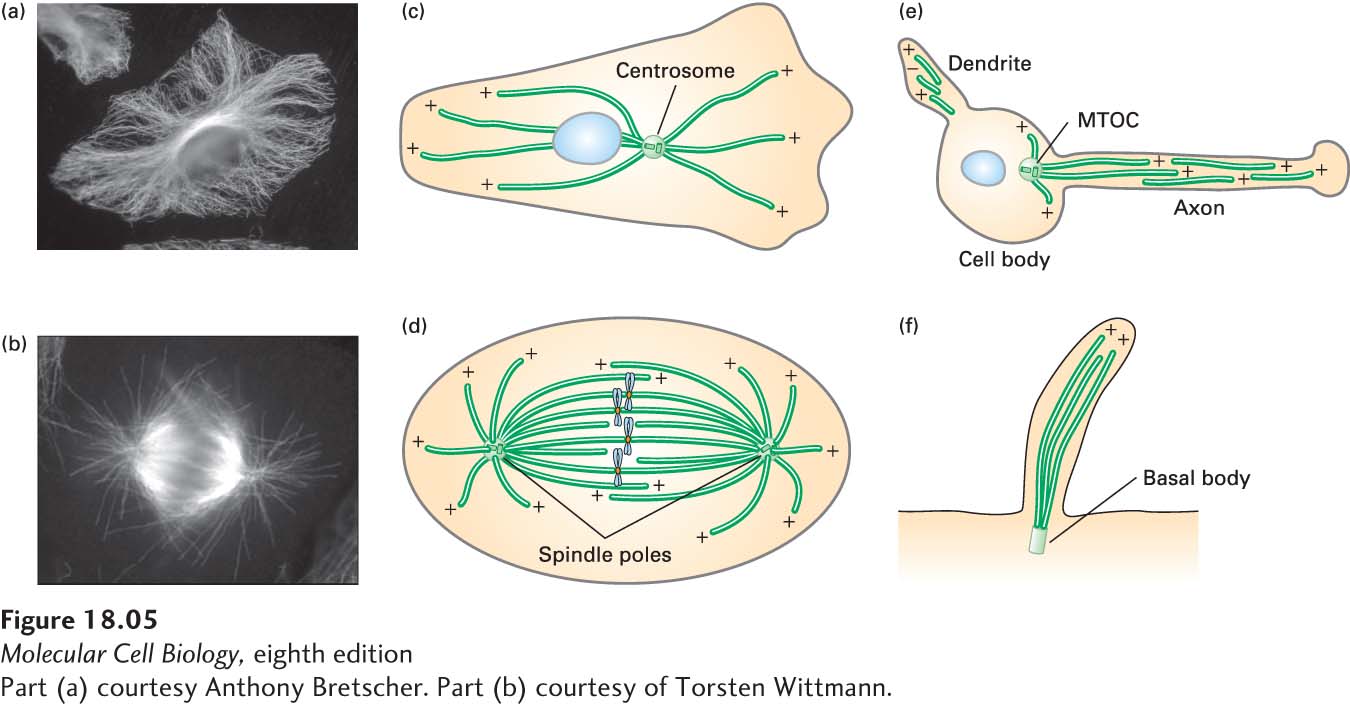
Once tubulin had been identified as the major structural component of microtubules, antibodies to tubulin could be generated and used in immunofluorescence microscopy to localize microtubules in cells (Figure 18-5a, b). This approach, coupled with electron microscopy, showed that microtubules are assembled from specific sites in the cell to generate many different configurations.
[Part (a) courtesy Anthony Bretscher. Part (b) courtesy of Torsten Wittmann.]
FIGURE 18-5Microtubules are assembled from microtubule organizing centers (MTOCs). (a–b) The distribution of microtubules in cultured cells, as seen by immunofluorescence microscopy using antibodies to tubulin, in an interphase cell (a) and a cell in mitosis (b). (c–f) Diagrams of the distributions of microtubules in various cells and structures. All of these microtubules are assembled from distinct MTOCs. (c) In an interphase cell, the MTOC is called a centrosome (the nucleus is indicated by a blue oval). (d) In a mitotic cell, the two MTOCs are called spindle poles (the chromosomes are shown in blue). (e) In a neuron, microtubules in both axons and dendrites are assembled from an MTOC in the cell body and then released from it. (f) The microtubules that make up the shaft of a cilium or flagellum are assembled from an MTOC known as a basal body. The polarity of microtubules is indicted by (+) and (−).
[Part (a) courtesy Anthony Bretscher. Part (b) courtesy of Torsten Wittmann.]
The nucleation phase of microtubule assembly is such an energetically unfavorable reaction that spontaneous nucleation does not play a significant role in microtubule assembly in vivo. Rather, all microtubules are nucleated from structures known as microtubule-organizing centers, or MTOCs. In most cases, the (−) end of the microtubule stays anchored in the MTOC while the (+) end extends away from it.
Page 825
There are several types of MTOCs. The centrosome is the main MTOC in animal cells. During interphase, the centrosome is generally located near the nucleus, producing an array of microtubules with their (+) ends radiating toward the cell periphery (Figure 18-5c). This radial array provides tracks for microtubule-based motor proteins to organize and transport membrane-bounded compartments, such as those constituting the secretory and endocytic pathways. During mitosis, cells completely reorganize their microtubules to form a bipolar spindle extending from two centrosomes, also known as spindle poles, that can accurately segregate copies of the duplicated chromosomes (Figure 18-5d). Neurons, in another example, have long processes called axons, in which organelles are transported in both directions along microtubules (Figure 18-5e). The microtubules in axons, which can be as long as 1 m, are not continuous and have been released from the MTOC, but nevertheless all have the same polarity. The microtubules in shorter processes, called dendrites, have mixed polarity, although the functional significance of this difference is not clear. In cilia and flagella (Figure 18-5f), microtubules are assembled from an MTOC called a basal body. As we mention later, plants do not have centrosomes and basal bodies, but use other mechanisms to nucleate the assembly of microtubules.
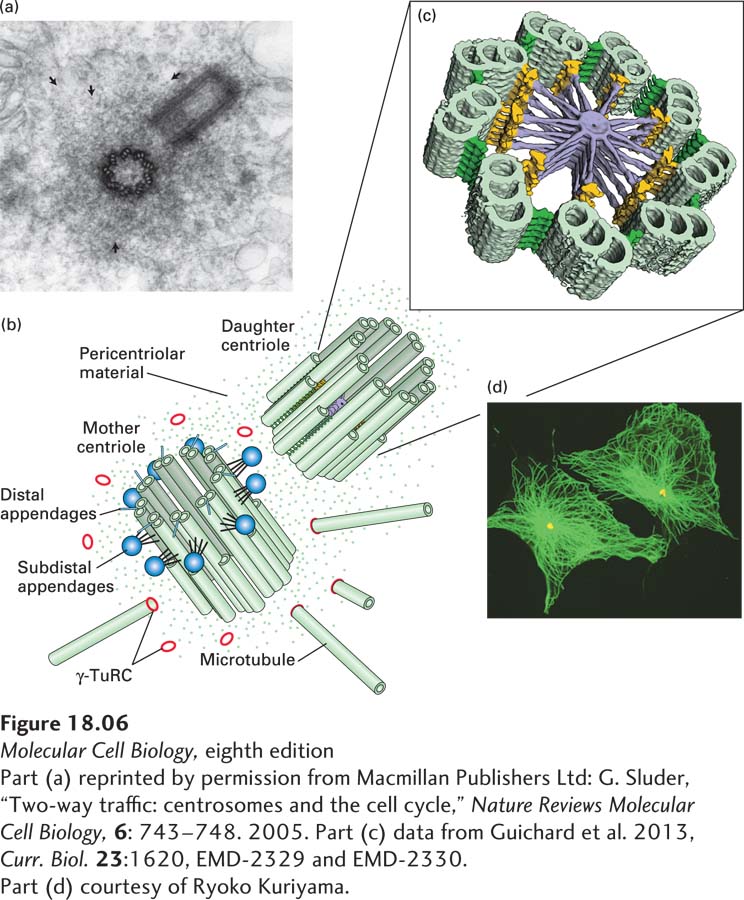
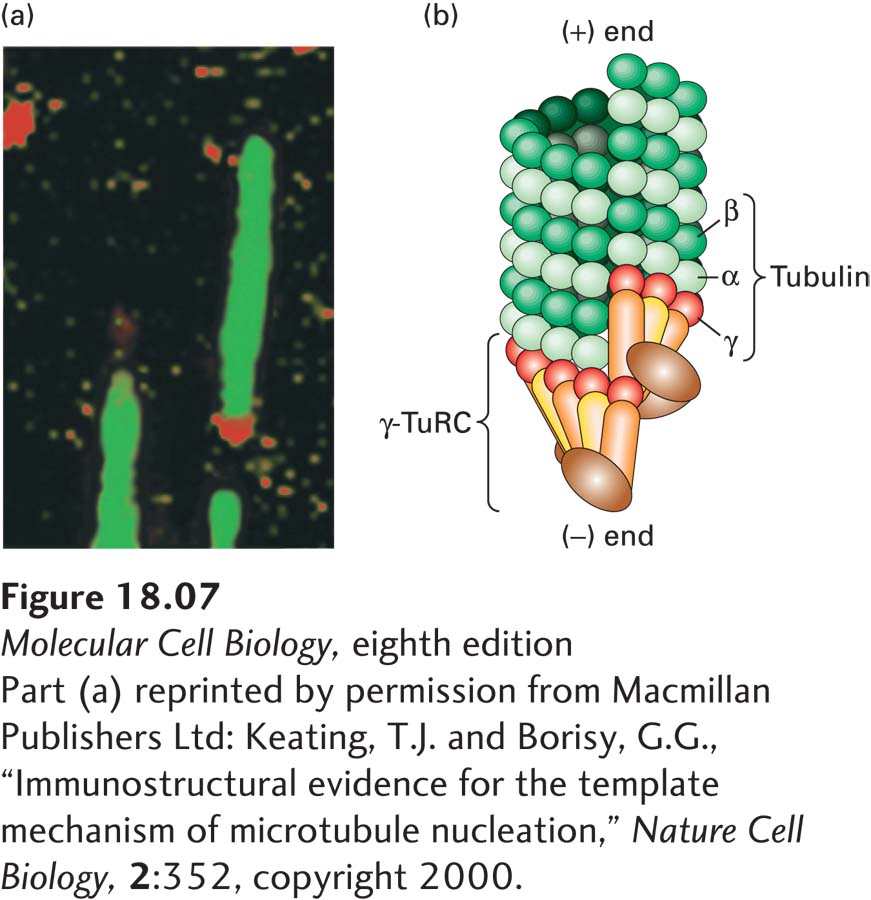
Electron microscopy shows that each centrosome in an animal cell consists of a pair of orthogonally arranged cylindrical centrioles surrounded by apparently amorphous material called pericentriolar material (Figure 18-6a). The centrioles, which are about 0.5 µm long and 0.2 µm in diameter, are highly organized and stable structures that consist of nine sets of triplet microtubules (Figure 18-6b). They are closely related in structure to the basal bodies found at the bases of cilia and flagella. It is not the centrioles themselves that nucleate the cytoplasmic microtubule array, but rather factors in the pericentriolar material. A critical component is the γ-tubulin ring complex (γ-TuRC) (Figure 18-6c and Figure 18-7), which is located in the pericentriolar material and consists of many copies of γ-tubulin associated with several other proteins. It is believed that γ-TuRC acts like a split-washer template to bind αβ-tubulin dimers for the formation of a new microtubule, whose (−) end is associated with γ-TuRC and whose (+) end is free for further assembly. In addition to nucleating the assembly of microtubules, centrosomes anchor and regulate the dynamics of the (−) ends of the microtubules, which are located there.
[Part (a) reprinted by permission from Macmillan Publishers Ltd: G. Sluder, “Two-way traffic: centrosomes and the cell cycle,” Nature Reviews Molecular Cell Biology,6: 743-748. 2005. Part (c) data from Guichard et al. 2013, Curr. Biol.23:1620, EMD-2329 and EMD-2330. Part (d) courtesy of Ryoko Kuriyama.]
FIGURE 18-6Structure of centrosomes. (a) Thin section of an animal-cell centrosome showing the two centrioles at right angles to each other, surrounded by pericentriolar material (arrows). (b) Diagram of a centrosome showing the mother and daughter centrioles, each of which consists of nine linked triplet microtubules, embedded in pericentriolar material that contains γ-TuRC nucleating structures. The mother centriole is distinct from the daughter as it has distal appendages (blue spheres). (c) A tomographic image of a section through a daughter centriole of the alga Chlamydomonas. The daughter centriole is believed to be templated by a nine-fold symmetric cartwheel structure that is later removed. (d) Immunofluorescence micrograph showing the microtubule array (green) in a cultured animal cell and the location of the MTOC, using an antibody to a centrosomal protein (yellow).
[Part (a) reprinted by permission from Macmillan Publishers Ltd: G. Sluder, “Two-way traffic: centrosomes and the cell cycle,” Nature Reviews Molecular Cell Biology,6: 743-748. 2005. Part (c) data from Guichard et al. 2013, Curr. Biol.23:1620, EMD-2329 and EMD-2330. Part (d) courtesy of Ryoko Kuriyama.]
[Part (a) reprinted by permission from Macmillan Publishers Ltd: Keating, T.J. and Borisy, G.G., “Immunostructural evidence for the template mechanism of microtubule nucleation,” Nature Cell Biology,2:352, copyright 2000.]
FIGURE 18-7The γ-tubulin ring complex (γ-TuRC) that nucleates microtubule assembly. (a) Immunofluorescence micrograph in which microtubules assembled in vitro are labeled green and a component of the γ-TuRC is labeled red, showing that it is located specifically at one end of the microtubule. (b) Model of how γ-TuRC nucleates assembly of a microtubule by forming a template corresponding to the (−) end.
[Part (a) reprinted by permission from Macmillan Publishers Ltd: Keating, T.J. and Borisy, G.G., “Immunostructural evidence for the template mechanism of microtubule nucleation,” Nature Cell Biology,2:352, copyright 2000.]
Basal bodies, which have a structure similar to that of a centriole, are the MTOCs found at the bases of cilia and flagella. The A and B tubules of their triplet microtubules provide a template for the assembly of the microtubules making up the core structure of cilia and flagella.
Recent work has uncovered an additional mechanism for the nucleation of microtubules in animal cells, also involving γ-TuRC. A protein complex called the augmin complex, consisting of eight polypeptides, can bind to the sides of existing microtubules, then recruit γ-TuRC and nucleate the assembly of new ones. As we discuss in a later section, the augmin complex contributes to microtubule assembly in the mitotic spindle.