Additional Mechanisms Contribute to Spindle Formation
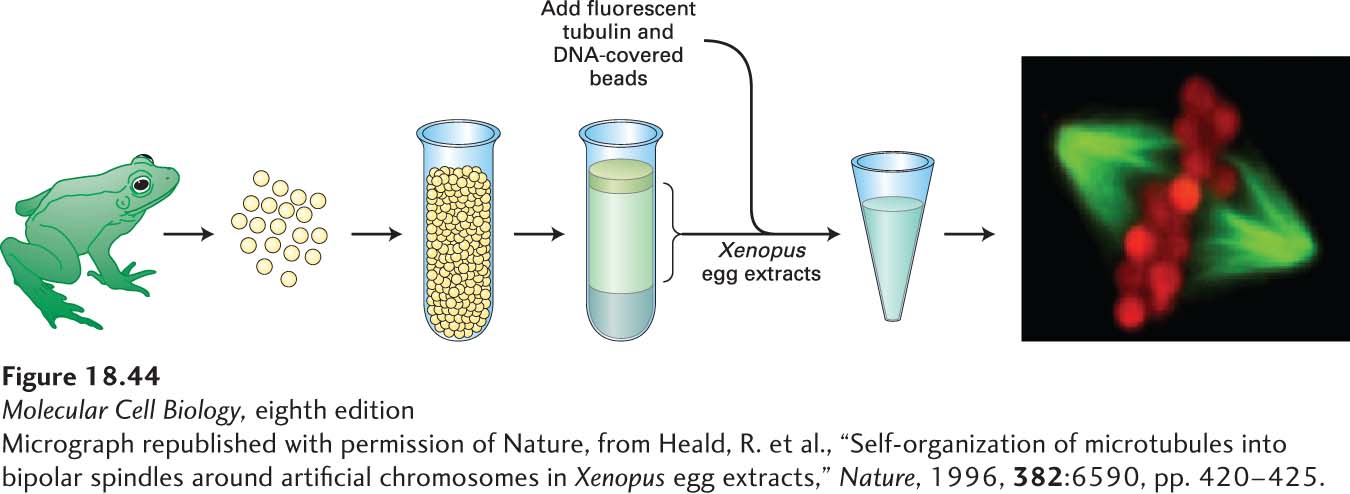
There are a number of cases in vivo in which spindles form in the absence of centrosomes, including plant-cell mitosis and animal-cell meiosis in females. This observation implies that nucleation of microtubules from centrosomes is not the only way in which a spindle can form. Studies exploiting mitotic extracts from frog eggs—extracts that do not contain centrosomes—show that the addition of beads covered with DNA is sufficient to assemble a relatively normal mitotic spindle (Figure 18-44). In this system, the beads induce the assembly of microtubules, and factors in the extract cooperate to make a spindle. One of the factors necessary for this reaction is cytoplasmic dynein, which is proposed to bind to two microtubules and migrate to their (−) ends, thereby drawing them together.
EXPERIMENTAL FIGURE 18-44Mitotic spindles can form in the absence of centrosomes. Centrosome-free extracts can be isolated from frog oocytes arrested in mitosis by centrifuging eggs to separate a soluble material from the organelles and yolk. When fluorescently labeled tubulin (green) is added to extracts of the soluble material together with beads covered with DNA (red), mitotic spindles spontaneously form around the beads from randomly nucleated microtubules. See Kinoshita et al., 2002, Trends Cell Biol.12:267–273, and Antonio et al., 2000, Cell102:425. [Micrograph republished with permission of Nature, from Heald, R. et al., “Self-organization of microtubules into bipolar spindles around artificial chromosomes in Xenopus egg extracts,” Nature, 1996, 382:6590, pp. 420-425.]
As mentioned in Section 18.1, a newly discovered γ-TuRC-associated complex, the augmin complex, also contributes microtubules to the mitotic spindle. In late prometaphase and metaphase, the augmin complex binds the sides of existing spindle microtubules to nucleate the assembly of additional microtubules having the same polarity as the mother microtubule. This activity contributes to both polar and kinetochore microtubule abundance in the spindle.
Recent studies have provided a model for how spindles can form in the absence of centrosomes. The ability of DNA-covered beads to induce the assembly of microtubules is dependent on Ran, the small GTPase that, in interphase cells, drives nuclear import and export (see Section 13.6). Recall that proteins destined for import into the nucleus associate with a nuclear import receptor, called an importin, that results in the complex passing from the cytosol through nuclear pores and into the nucleoplasm. The level of Ran·GTP is high in the nucleus because its activator, the guanine nucleotide exchange factor for Ran (Ran-GEF), is localized there by binding to chromatin. Ran⋅GTP binds to the importin, changing its conformation to release the transported protein, and then the importin-Ran⋅GTP complex leaves the nucleus to return to the cytosol through a nuclear pore. After nuclear envelope breakdown in mitosis, because the Ran-GEF is associated with chromatin, there is a gradient of Ran⋅GTP around chromosomes. This Ran⋅GTP releases a protein called TPX from importin. TPX binds the augmin complex and γ-TuRC to drive the assembly of microtubules in the vicinity of the chromatin, which can be used to make the spindle. In spindles with centrosomes, this pathway also exists to enhance the number of kinetochore and polar microtubules. It involves an association of the TPX–augmin–γ-TuRC complex with the side of an existing microtubule to nucleate assembly of new ones at a shallow angle to the mother microtubule, and thereby enhance the density of microtubules with the same polarity.