Lamins Line the Inner Nuclear Envelope To Provide Organization and Rigidity to the Nucleus
The most widespread IF proteins are the class V proteins, the lamins. The lamins are the progenitors of all IF proteins, from which the cytoplasmic IF proteins arose by gene duplication and mutation. They are the major components of a two-dimensional meshwork called the nuclear lamina that lies between the nuclear envelope and the chromatin of the nucleus (Figure 18-51a). In humans, three genes encode lamins: one alternatively spliced gene encodes lamins A and C, and two other genes encode lamins B1 and B2. The B-type lamins appear to be the primordial lamin proteins and are expressed in essentially all cells, whereas lamins A and C are developmentally regulated. B lamins are post-translationally prenylated (see p. 288), which helps them associate with the inner nuclear envelope membrane. Lamin proteins contain the coiled-coil regions characteristic of intermediate filaments that is needed for dimerization, but also have a nuclear localization sequence that targets them to the nucleus as well as a conserved immunoglobulin-like fold (see Figure 18-48d).
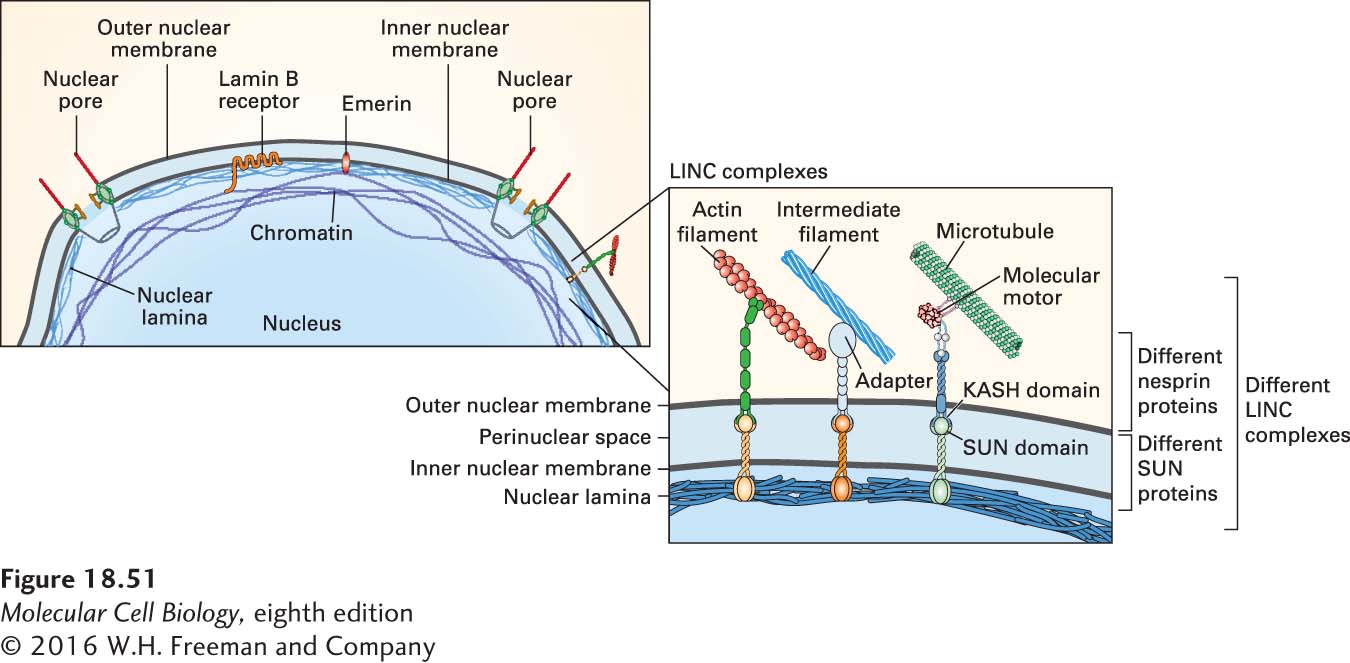
FIGURE 18-51The nuclear lamina is attached to chromatin and through LINC complexes to the cytoskeleton. (a) Diagram of part of a nucleus, showing the association of the lamin-containing nuclear lamina with chromatin and, through the two membranes of the nucleus, with the cytoskeleton. Proteins such as the membrane-associated lamin B receptor and emerin tether the lamin intermediate filaments to the inner nuclear membrane. Lamins are also tethered to the nuclear membrane by the prenylation of lamin B (not shown). Diverse linkages, called LINC complexes, attach the lamins through the two nuclear membranes to the cytoskeleton. (b) A LINC complex consists of a SUN domain–containing protein that interacts with lamins and extends across the inner nuclear membrane, and a KASH domain–containing protein that interacts with a SUN domain–containing protein in the perinuclear space and crosses the outer nuclear membrane to interact with components of the cytoskeleton. See B. Burke and C. L. Stewart, 2013, Nat. Rev. Mol. Cell Biol.14:13.
Cells that are subject to mechanical stress have a nuclear lamin meshwork to maintain the integrity of the nucleus; cells with strong cell walls, such as plant and fungal cells, do not have such a meshwork. The lamin meshwork therefore provides both strength and support to the inner surface of the nuclear membrane. In fact, cells regulate the level of lamin A to match the stiffness of the tissue they are in. Thus neutrophils, for example, which need to move through thin capillaries and migrate through tight interstitial spaces, have a highly lobulated nucleus as a result of very low levels of lamin A. If they had large, rigid nuclei that were resistant to deformation, they might have trouble squeezing through the small spaces found in the extracellular matrix.
To provide rigidity, the lamin meshwork is associated with chromatin on one side and attached to the cytoskeleton on the other. Some proteins embedded in the inner nuclear membrane, such as the lamin-B receptor and emerin, can bind both chromatin-associated proteins and lamins (see Figure 18-51a). Interestingly, transcriptionally silent regions of the genome are preferentially associated with lamins, and recent evidence suggests that lamins play a role in genome organization and DNA repair. Attachment to the cytoskeleton through both the inner and outer nuclear membranes involves proteins with so-called SUN and KASH domains. The SUN domain proteins are synthesized on the endoplasmic reticulum as transmembrane proteins with their SUN domain in the lumen of the endoplasmic reticulum, and sorting signals in their cytoplasmic domain target them to the outer nuclear membrane, which is continuous with the endoplasmic reticulum. They are then transported as membrane proteins through nuclear pores and, upon reaching the inner nuclear membrane, associate with the nuclear lamina (Figure 18-51b). Nesprins are KASH domain–containing transmembrane proteins of the outer nuclear membrane oriented so that the KASH domain can associate with the SUN domain of another protein in the perinuclear space. The nesprins, in turn, associate, either directly or through adapters, with intermediate filaments, actin filaments, and microtubules, thereby providing physical linkage of the nucleus to the cytoskeleton (see Figure 18-51b). These attachments are used to move the nucleus to the correct location in a cell as well as transport it, for example, in the long processes of the vertebrate neuroepithelium.