Cells Are Irreversibly Committed to Division at a Cell Cycle Point Called START or the Restriction Point
In most eukaryotic cells, the key decision that determines whether or not a cell will divide is whether or not to enter S phase. In most cases, once a cell has become committed to entering the cell cycle, it must complete it. The budding yeast Saccharomyces cerevisiae regulates its proliferation in this manner, and much of our current understanding of the molecular mechanisms controlling entry into the cell cycle originated with genetic studies of S. cerevisiae.
Recall that S. cerevisiae exists in two different mating types, a and α (see Fig. 1-23). These two haploid cell types can mate to form an a/α diploid cell. Mating is initiated by secreted pheromones and ultimately results in the fusion of two cells of opposite mating type. It is essential for successful mating that both mating partners be arrested in G1. Thus, in addition to inducing genes critical for mating, the pheromones cause cells to arrest in G1. If cells are in G1, addition of pheromones will keep cells arrested in G1, but once a cell has committed to divide, it must complete a whole cell cycle before it can be arrested in G1 again by pheromones. The point in late G1 when S. cerevisiae cells become irrevocably committed to entering and traversing the entire cell cycle even when pheromones are present is called START. A similar transition point, called the restriction point, exists in mammalian cells at which cells become refractory to growth factor signals and proliferation inhibitor signals.
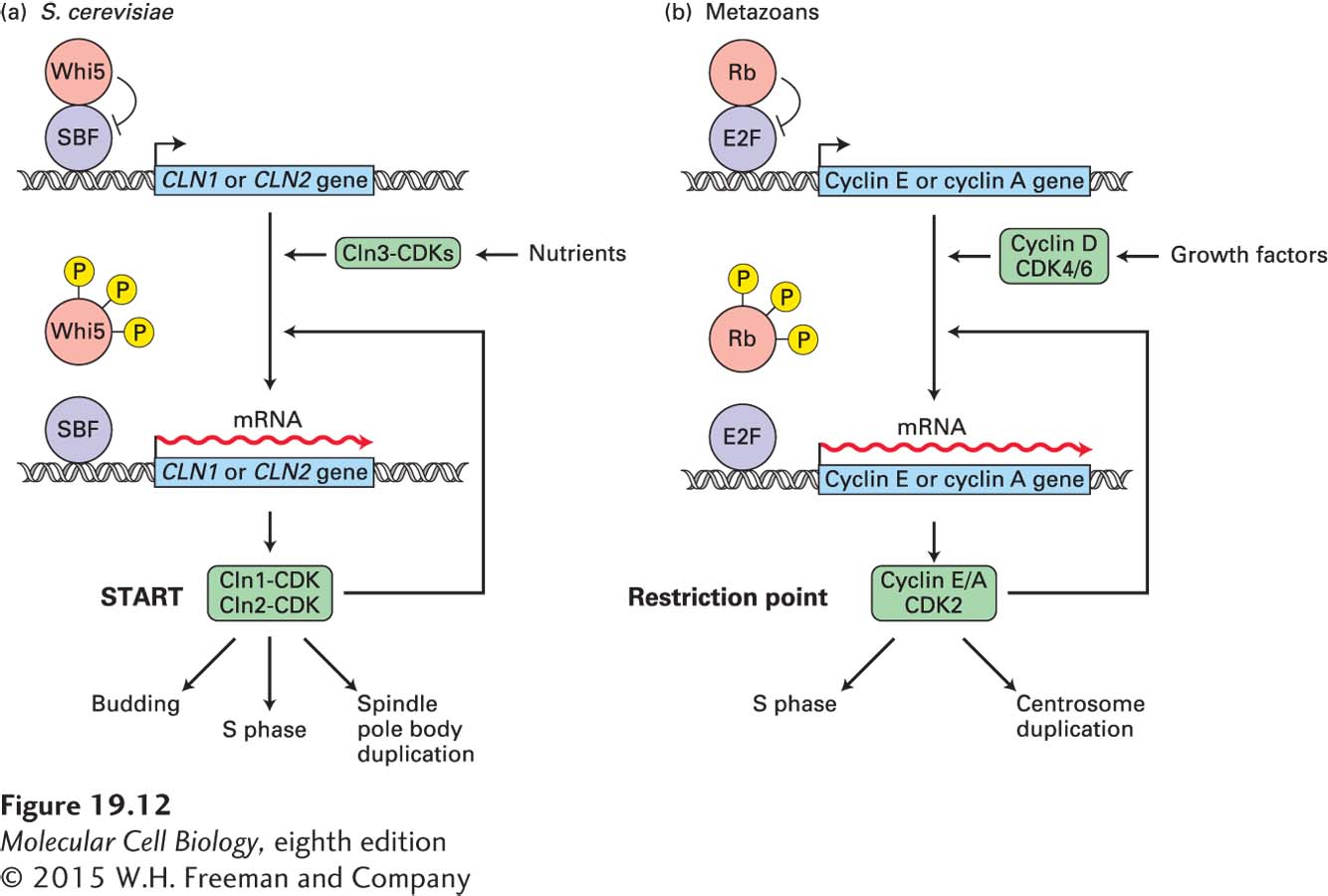
CDK activity is essential for entry into S phase. This was first realized in budding yeast, in which temperature-sensitive CDK mutants arrest in G1, failing to form a bud and to initiate DNA replication (budding yeast has only one CDK that triggers all cell cycle transitions and is known as CDC28). We now know that a CDK cascade triggers entry into the cell cycle. G1 cyclin-CDK complexes stimulate the formation of G1/S phase cyclin-CDKs, which then initiate bud formation, centrosome duplication, and DNA replication. In yeast, the G1 cyclin gene is called CLN3 (Figure 19-12a). Its mRNA is produced at a nearly constant level throughout the cell cycle, but its translation is regulated in response to nutrient levels and, as we will see shortly, it is a linchpin in coupling cell cycle entry to nutrient signals. Once sufficient Cln3 is synthesized from its mRNA, Cln3-CDK complexes phosphorylate and inactivate the transcriptional repressor Whi5. Phosphorylation of Whi5 promotes its export out of the nucleus, allowing the transcription factor complex SBF to induce transcription of the G1/S phase cyclin genes CLN1 and CLN2 as well as other genes important for DNA replication. Once produced, Cln1/2-CDKs contribute to further Whi5 phosphorylation. This positive feedback loop ensures the rapid accumulation of G1/S phase cyclin-CDKs. The point in the cell cycle at which 50 percent of Whi5 has exited the nucleus is the point when cells are irreversibly committed to division. It is the molecular definition of START. Cln1/2-CDKs then trigger bud formation, entry into S phase, and the duplication of the centrosome (also known as the spindle pole body in yeast), which later in the cell cycle will organize the mitotic spindle.
FIGURE 19-12Control of the G1–S phase transition. (a) In budding yeast, Cln3-CDK activity rises during G1 and is controlled by nutrient availability. Once sufficiently active, the kinase phosphorylates the transcriptional repressor Whi5, promoting its export from the nucleus. This causes the transcription factor complex SBF to induce the transcription of the G1/S phase cyclin genes CLN1 and CLN2 and of other genes whose products are needed for DNA replication. G1/S phase CDKs further phosphorylate Whi5, promoting further CLN1 and CLN2 transcription. Once sufficiently high levels of G1/S phase CDKs have been produced, START is traversed. Cells enter the cell cycle: they initiate DNA replication, bud formation, and spindle pole body duplication. (b) In vertebrates, G1 CDK activity rises during G1 and is stimulated by the presence of growth factors. When signaling from growth factors is sustained, the resulting cyclin D–CDK4/6 complexes begin phosphorylating Rb, releasing some E2F, which stimulates transcription of the genes encoding cyclin E, CDK2, and E2F itself. The cyclin E–CDK2 complexes further phosphorylate Rb, resulting in a positive feedback loop that leads to a rapid rise in the expression and activity of both E2F and cyclin E–CDK2. Once G1/S phase CDKs are sufficiently high, cells pass through the restriction point. They commence DNA replication and centrosome duplication.