Duplicated DNA Strands Become Linked During Replication
During S phase, as chromosomes are duplicated to form sister chromatids, they become tethered to each other by protein links. The linkages between sister chromatids established during S phase will be essential for their accurate segregation during mitosis.
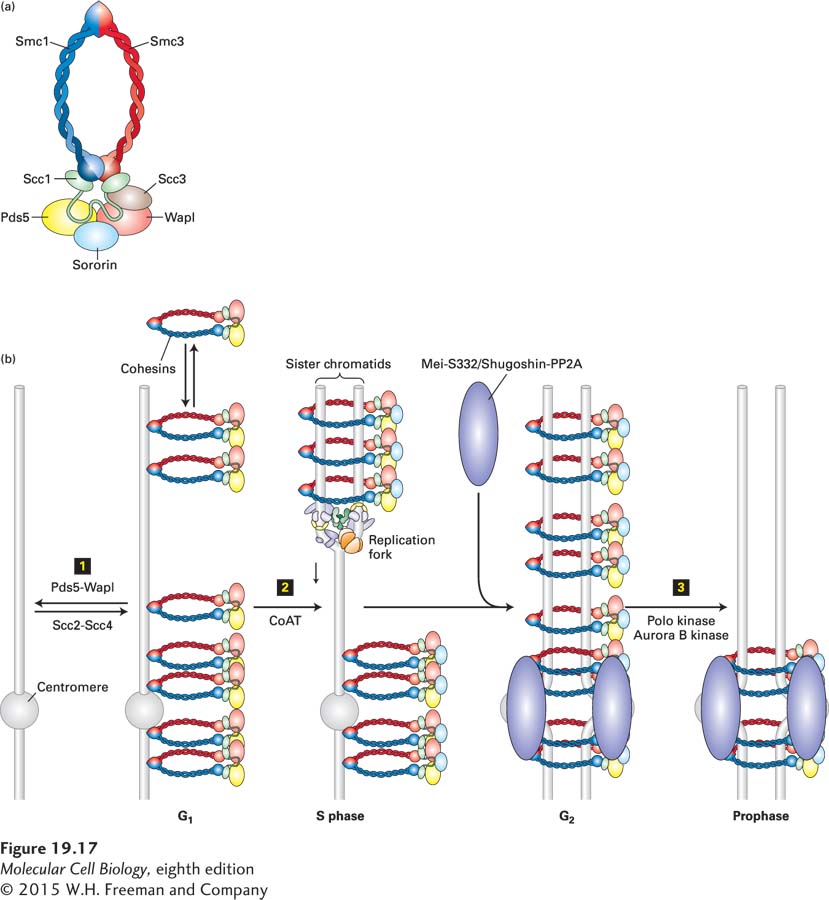
The protein complexes that establish these linkages between sister chromatids are called cohesins. They are composed of four subunits: Smc1, Smc3, Scc1 (sometimes called Rad21), and Scc3 (Figure 19-17a). Smc1 and Smc3 are members of the SMC protein family, which is characterized by long coiled-coil domains that are flanked by a globular domain containing ATPase activity. The ATPase domains interact with Scc1 and Scc3 and, together, form a ring structure. These rings of cohesin embrace one or both copies of the replicated DNA. When cohesins are inactivated, sister chromatids do not associate properly with each other.
Page 894
FIGURE 19-17Model for establishment of cohesin linkage of sister chromatids. Cohesin complexes form rings that link sister chromatids by embracing the two sister DNA molecules. (a) Schematic structure of the cohesin complex. (b) Mechanism whereby cohesins are loaded onto DNA and acquire their cohesive properties. Step 1: Cohesins are loaded onto chromosomes during G1 by the cohesin-loading complex Scc2-Scc4, but they do not possess cohesive properties (indicated as cohesins laterally associated with chromosomes). In this state, cohesins are dynamic and can dissociate from the DNA with the help of the Pds5-Wapl complex, which associates with cohesins. Step 2: Concomitant with DNA replication, closely behind the replication fork, cohesins are converted into cohesive molecules, able to hold sister chromatids together (indicated as cohesin rings encircling the replicated sister chromatids), through acetylation of Smc3 by cohesin acetyltransferases (CoATs). Acetylation is accompanied by the binding of sororin to cohesin, which helps stabilize cohesins on chromosomes. During G2, sister chromatids are replicated and linked along their entire length by cohesins. During this time, the Mei-S332/Shugoshin proteins recruit the protein phosphatase 2A (PP2A) to centromeric regions. Step 3: In vertebrate cells, cohesins are released from chromosome arms during prophase and early metaphase by the action of the Pds5-Wapl complex and phosphorylation of cohesins by Polo kinase and Aurora B kinase. By the end of metaphase, cohesins are retained only in the region of the centromere, where Mei-S332/Shugoshin prevents cohesin phosphorylation, and hence dissociation, by recruiting PP2A.
Cohesin-mediated cohesion between sister chromatids is established by a two-step process and is tightly tied to DNA replication. Cohesins associate with chromosomes during G1; this association requires the cohesin loading factors Scc2 and Scc4 (Figure 19-17b, step 1). In this phase of the cell cycle, cohesins are quite dynamic on chromosomes. They are continuously unloaded by a cohesin-associated complex composed of the Pds5 and Wapl proteins. This interphase dynamicity of cohesins is likely to be important for their role in regulating interphase chromatin structure and gene expression. Cohesins acquire their cohesive properties during DNA replication. The two duplicated DNA strands become entrapped within the cohesin rings as replication forks replicate the DNA (Figure 19-17b, step 2). Converting DNA-bound G1 cohesins into cohesive complexes requires acetylation of the Smc3 subunit by cohesin acetyl transferases (CoATs). This acetylation prevents unloading of cohesins by Pds5-Wapl, thereby stabilizing the cohesins on chromosomes. In vertebrates, this stabilization of cohesins requires the cohesin-associated factor sororin (Figure 19-17b, step 2). As we will see in Section 19.6, cohesins are essential for accurate attachment of the replicated sister chromatids to the mitotic spindle and for their segregation during mitosis. Cells lacking cohesins or the factors that load them onto chromosomes segregate chromosomes randomly.
Page 895
Cohesins are not only critical for establishing linkages between replicated DNA molecules, and hence for their accurate segregation during mitosis, but also regulate gene expression. The functions of cohesins in this process appear to be diverse. In some instances, cohesins promote gene expression, whereas in others they restrain it. The mechanism whereby cohesins accomplish gene expression control is, however, the same in both cases: cohesins promote chromatin loop formation, thereby bringing enhancer or repressive elements close to the transcriptional start site. Defects of cohesin’s gene expression regulation function are the cause of a group of diseases collectively called cohesinopathies. In these diseases, mutations in cohesin subunits or cohesin loading factors disrupt the expression of genes that are critical for development, causing limb and craniofacial abnormalities and intellectual disabilities. Cohesin’s sister chromatid cohesion function, however, appears intact in these diseases. In contrast, as we will see in Section 19.8, defects in cohesin’s cohesion function during meiosis cause miscarriages and intellectual disabilities.