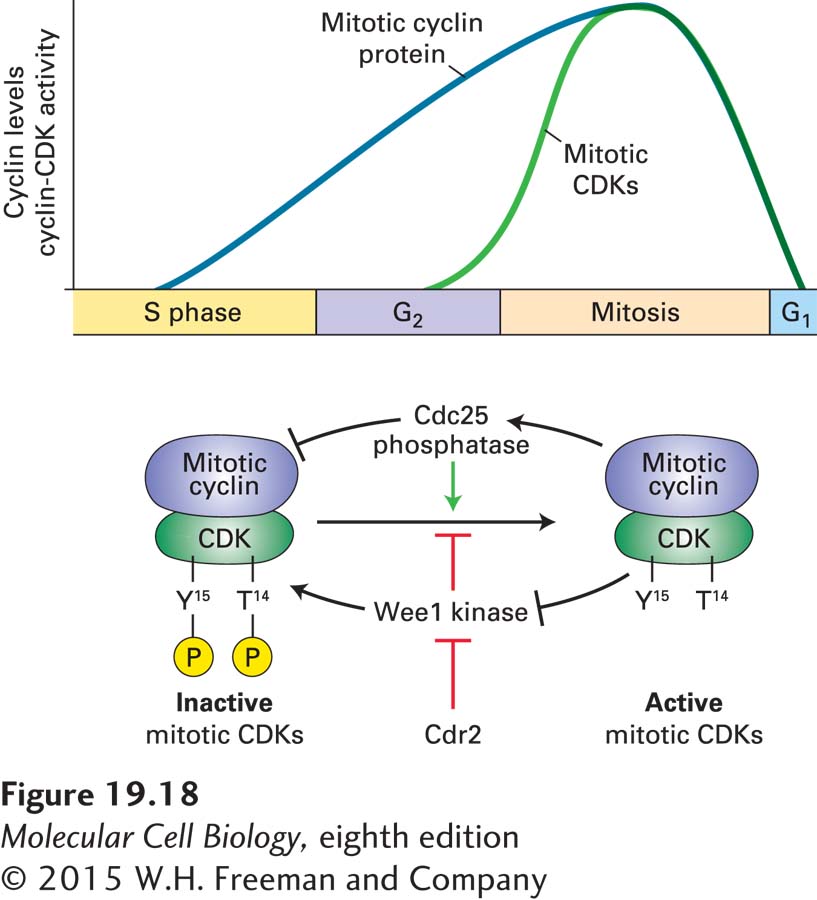
Precipitous Activation of Mitotic CDKs Initiates Mitosis
Mitotic cyclin-CDKs initiate mitosis. Whereas levels of the catalytic CDK subunit are constant throughout the cell cycle, mitotic cyclins gradually accumulate during S phase. Most eukaryotes contain multiple mitotic cyclins, which for historical reasons are subdivided into the cyclin A and cyclin B families. As they assemble, mitotic CDK complexes are maintained in an inactive state through inhibitory phosphorylation of the CDK subunit. Recall from Section 19.3 that two highly conserved tyrosine and threonine residues in mammalian CDKs are subject to regulated phosphorylation. In CDK1, the mitotic CDK, phosphorylation of tyrosine 15 and threonine 14 |maintains mitotic cyclin-CDK complexes in an inactivate state. The phosphorylation state of T14 and Y15 is controlled by a dual-specificity protein kinase known as Wee1 and a dual-specificity phosphatase, Cdc25 (Figure 19-18). Such kinases and phosphatases can phosphorylate and dephosphorylate both serines/threonines and tyrosines, respectively. The regulation of mitotic CDKs by these activities underlies the abrupt activation of their kinase activity at the G2–M phase transition and explains the observation that although mitotic cyclins gradually accumulate during S phase and G2, mitotic CDKs are not active until cells enter mitosis.
FIGURE 19-18Phosphorylation of the CDK subunit restrains mitotic CDK activity during S phase and G2. Mitotic cyclins are synthesized during S phase and G2 and bind to CDK1. However, the cyclin-CDK complex is not active because threonine 14 and tyrosine 15 of the CDK1 subunit are phosphorylated by the protein kinase Wee1. Once DNA replication has been completed, the protein phosphatase Cdc25 is activated and dephosphorylates CDK1. Active mitotic CDKs further stimulate Cdc25. At the same time, mitotic CDKs inhibit Wee1, the protein kinase that places the inhibitory phosphorylation on the CDK subunit. Ongoing DNA replication inhibits Cdc25 activity. How Cdc25 is initially activated upon completion of DNA replication to put these feedback loops in motion is not yet known. Cell size also affects this regulatory loop. Once cells reach the appropriate size, Cdr2 inhibits Wee1, allowing Cdc25 to activate mitotic CDKs.
Studies in the fission yeast Schizosaccharomyces pombe unraveled the mechanisms that lead to the precipitous activation of mitotic CDKs during G2. The dual-specificity protein kinase Wee1 phosphorylates CDKs on the inhibitory tyrosine 15. (Threonine 14 is not phosphorylated in S. pombe CDK1.) Yeast cells with a defective wee1+ gene activate mitotic CDKs prematurely and hence experience premature entry into mitosis. These wee1 mutants not only enter mitosis prematurely, but are also smaller. This is because unlike most other eukaryotes, which coordinate cell size and cell division during G1, this coordination occurs during G2 in fission yeast. Fission yeast cells carrying a CDK1 mutation in which the tyrosine 15 residue is replaced by phenylalanine (which is structurally similar to tyrosine but cannot be phosphorylated) show the same premature mitotic CDK activation and entry into mitosis. Fission yeast cells carrying mutations in the cdc25+ gene arrest in G2, indicating that the phosphatase that opposes Wee1 is essential for entry into mitosis.
Vertebrates contain multiple Wee1 protein kinases and multiple Cdc25 phosphatases, which collaborate to control not only mitotic CDK activity but also the activity of G1/S phase CDKs. One member of the Cdc25 family of phosphatases, Cdc25A, is activated in late G1. It removes the inhibitory phosphorylation on tyrosine 15 of the G1/S phase CDKs and S phase CDK catalytic subunit to activate the kinases. Another family member, Cdc25C, is active during G2 and removes the inhibitory phosphorylation on mitotic CDKs.
Activation of mitotic CDKs is the consequence of rapid inactivation of Wee1 and activation of Cdc25. Central to this rapid transition are feedback loops, whereby mitotic CDKs activate Cdc25 and inactivate Wee1 (see Figure 19-18). Phosphorylation of Cdc25 by mitotic CDKs stimulates its phosphatase activity; phosphorylation of Wee1 by mitotic CDKs inhibits its kinase activity. Ongoing DNA replication inhibits Cdc25 activity. A critical question that we know little about is how this positive feedback loop is started once DNA replication has been completed. CDKs that function earlier in the cell cycle have been suggested to start the positive feedback loop.
Although it is not yet known how the precipitous activation of mitotic CDKs is initiated, it is clear that once active, these protein kinases set in motion all the events necessary to ready the cell for chromosome segregation. The activation of mitotic CDKs is associated with changes in the subcellular localization of these kinases. Mitotic CDKs initially associate with centrosomes, where they are thought to facilitate centrosome maturation. They then enter the nucleus, where they bring about chromosome condensation and nuclear envelope breakdown. In what follows, we discuss how the mitotic CDKs accomplish the coordinated execution of mitosis.
Page 897
During DNA replication initiation, S phase CDKs work together with DDK to promote MCM helicase activation. In a similar manner, mitotic CDKs collaborate with other protein kinases to bring about the mitotic events. The Polo kinase family is critical for formation of the mitotic spindle as well as for chromosome segregation. The Aurora kinase family plays key roles in mitotic spindle formation and in ensuring that chromosomes attach to the mitotic spindle in the correct way so that they are segregated accurately during mitosis. Their contributions to the various mitotic events will also be discussed.