Several Key Features Distinguish Meiosis from Mitosis
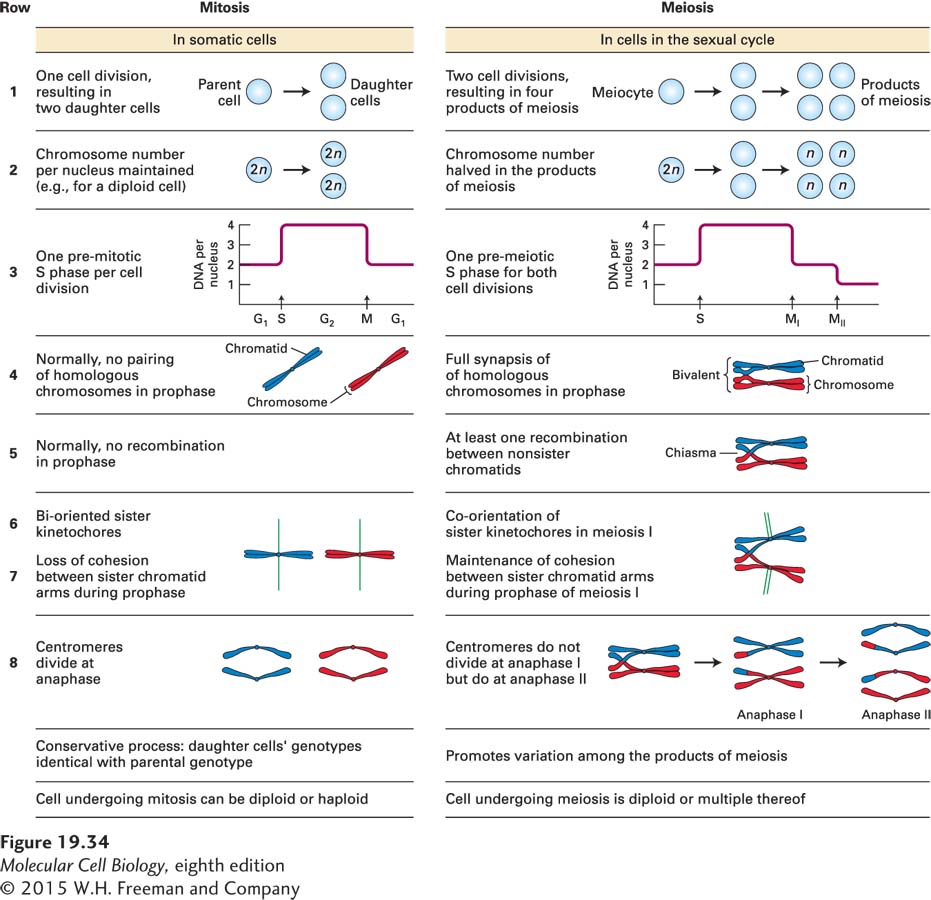
The meiotic divisions differ in several fundamental aspects from the mitotic divisions. These differences are summarized in Figure 19-34. During the meiotic cell division, a single round of DNA replication is followed by two cycles of cell division, termed meiosis I and meiosis II (Figure 19-35). Meiosis II resembles mitosis in that sister chromatids are segregated. However, meiosis I is very different. During this division, homologous chromosomes—the chromosome inherited from your mother and the same chromosome inherited from your father—are segregated. This unusual chromosome segregation requires three meiosis-specific modifications to the chromosome segregation machinery. In what follows, we discuss these modifications and explain why they are needed.
FIGURE 19-34Comparison of the main features of mitosis and meiosis.
[Photos courtesy of Ed Reschke/Peter Arnold, Inc./Photolibrary/Getty Images.]
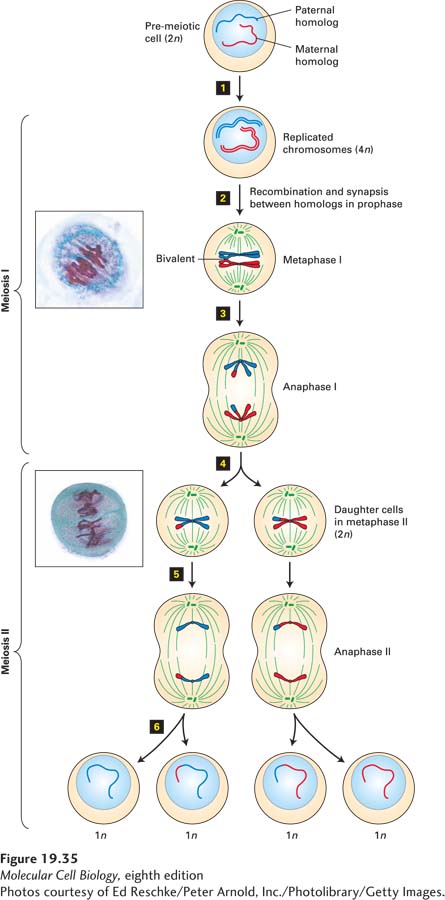
Figure 19-35Meiosis. Pre-meiotic cells have two copies of each chromosome (2n), one derived from the paternal parent and one from the maternal parent. For simplicity, the paternal and maternal homologs of only one chromosome are diagrammed. Step 1: All chromosomes are replicated during S phase before the first meiotic division, giving a 4n chromosomal complement. Cohesin complexes (not shown) link the sister chromatids composing each replicated chromosome along their full lengths. Step 2: As chromosomes condense during the first meiotic prophase, the replicated homologs pair and undergo homologous recombination, leading to at least one crossover event. At metaphase I, shown here, both chromatids of one chromosome associate with microtubules emanating from one spindle pole, but each member of a homologous chromosome pair associates with microtubules emanating from opposite poles. Step 3: During anaphase of meiosis I, the homologous chromosomes, each consisting of two chromatids, are pulled to opposite spindle poles. Step 4: Cytokinesis yields two daughter cells (now 2n), which enter meiosis II without undergoing DNA replication. At metaphase of meiosis II, shown here, the sister chromatids associate with spindle microtubules from opposite spindle poles, as they do in mitosis. Steps 5 and 6: Segregation of sister chromatids to opposite spindle poles during anaphase of meiosis II, followed by cytokinesis, generates haploid gametes (1n) containing one copy of each chromosome. Micrographs on the left show meiotic metaphase I and metaphase II in developing gametes from Lilium (lily) ovules. Chromosomes are aligned at the metaphase plate.
[Photos courtesy of Ed Reschke/Peter Arnold, Inc./Photolibrary/Getty Images.]
The tension-based sensing mechanism responsible for accurately attaching chromosomes to the spindle during mitosis is also responsible for segregating chromosomes during meiosis I. Thus homologous chromosomes must be linked so that this tension-based mechanism can function. Homologous recombination between homologous chromosomes creates these linkages (see Figure 19-34). The molecular mechanisms of homologous recombination are discussed in detail in Chapter 4. Here we restrict our discussion to the importance of homologous recombination to successful meiotic divisions.
In G2 and prophase of meiosis I, the two replicated chromatids of each chromosome are linked together by cohesin complexes along the full length of the chromosome arms, just as they are following DNA replication in a mitotic cell cycle. In prophase of meiosis I, homologous chromosomes (i.e., the maternal and paternal chromosome 1, the maternal and paternal chromosome 2, etc.) pair with each other and undergo homologous recombination. Significantly, at least one recombination event occurs between a maternal and a paternal chromosome. The crossing over of chromatids produced by recombination can be observed microscopically in the first meiotic prophase and metaphase as structures called chiasmata (singular, chiasma). In contrast, no pairing between homologous chromosomes occurs during mitosis, and recombination between nonsister chromatids is rare. Concomitant with homologous recombination, homologous chromosomes associate with each other in a process known as synapsis. In most organisms, this synapsis is mediated by a proteinaceous complex known as the synaptonemal complex (SC). Homologous chromosomes linked through chiasmata are called bivalents (see Figure 19-34). The chiasmata and the cohesin molecules distal to them now provide the resistance to the pulling force exerted by microtubules on the metaphase I spindle (see Figure 19-35).
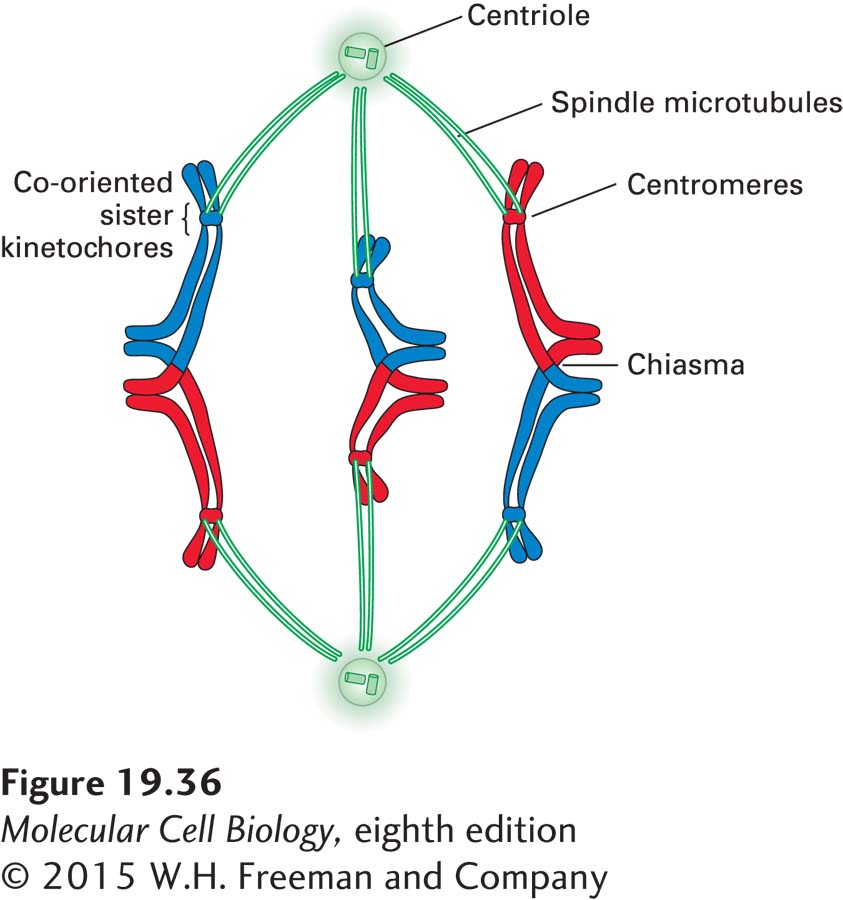
The recombination between homologous chromosomes that occurs in prophase of meiosis I has at least two functional consequences: First, it connects homologous chromosomes during meiosis I metaphase. Second, it contributes to genetic diversity among individuals of a species by ensuring new combinations of alleles in different individuals. (Note, however, that genetic diversity primarily arises from the independent reassortment of maternal and paternal homologs during the meiotic divisions.) The homologs, now connected through at least one chiasma, must align on the spindle in metaphase of meiosis I so that maternal and paternal chromosomes are segregated away from each other during anaphase of meiosis I. This requires that the kinetochores of sister chromatids attach to microtubules emanating from the same spindle pole rather than from opposite spindle poles, as in mitosis (Figure 19-36). Sister chromatids attached in this way are said to be co-oriented. However, the kinetochores of the maternal and paternal chromosomes of each bivalent attach to spindle microtubules from opposite spindle poles; they are bi-oriented.
Figure 19-36Chiasmata and cohesins distal to them link homologous chromosomes in meiosis I metaphase. Connections between chromosomes during meiosis I are most easily visualized in organisms with acrocentric centromeres, such as the grasshopper. The kinetochores at the centromeres of sister chromatids attach to spindle microtubules emanating from the same spindle pole, with the kinetochores of the maternal (red) and paternal (blue) chromosomes attaching to spindle microtubules from opposite spindle poles. The maternal and paternal chromosomes are attached to each other by chiasmata, which are formed by recombination between them, and by the cohesion between sister chromatid arms that persists until metaphase I. Note that elimination of cohesion between sister chromatid arms is all that is required for the homologous chromosomes to separate at anaphase. See L. V. Paliulis and R. B. Nicklas, 2000, J. Cell Biol.150:1223.
Page 913
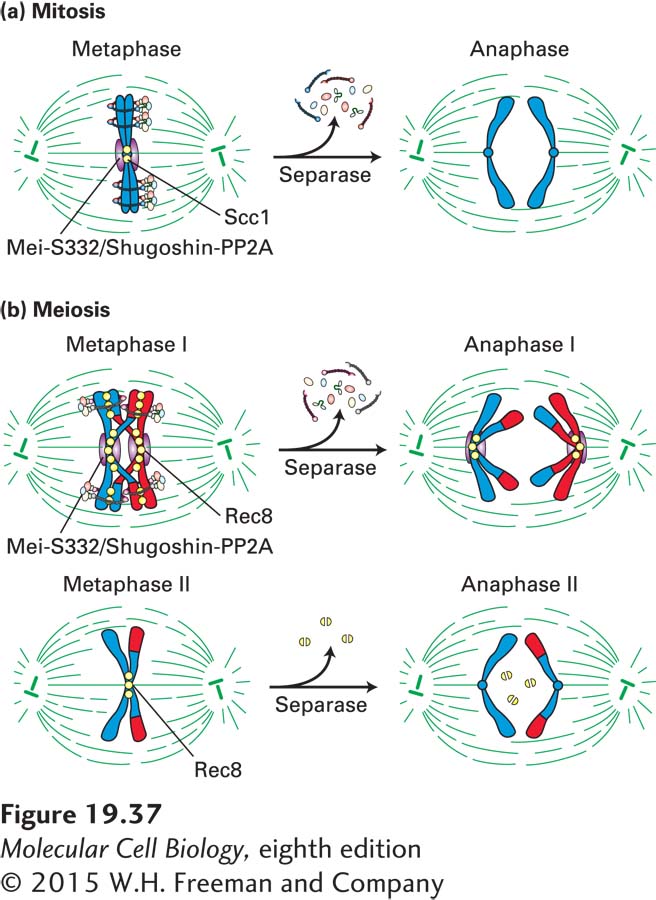
Finally, to facilitate two consecutive chromosome segregation phases, cohesins must be lost from chromosomes in a stepwise manner. Recall that during mitosis, all cohesins are lost by the onset of anaphase (Figure 19-37a). In contrast, during meiosis, cohesins are lost from chromosome arms by the end of meiosis I, but a pool of cohesins around kinetochores is protected from removal (Figure 19-37b). This pool of cohesins persists throughout meiosis I, but is removed at the onset of anaphase II. As we will see next, loss of cohesins from chromosome arms is required for homologous chromosomes to segregate away from each other during meiosis I.
Figure 19-37Cohesin function during mitosis and meiosis. (a) During mitosis, sister chromatids generated by DNA replication in S phase are initially linked by cohesin complexes along the length of the chromatids. During chromosome condensation, cohesin complexes (yellow) become restricted to the region of the centromere at metaphase. Mei-S332/Shugoshin (purple) recruits PP2A to centromeres, where they antagonize Polo kinase and Aurora B, preventing the dissociation of cohesins from centromeric regions. Dissociation of Mei-S332/Shugoshin from centromeres and activation of separase leads to removal of cohesins at the centromere. Sister chromatids now separate, marking the onset of anaphase. (b) In prophase of meiosis I, maternal and paternal chromatids establish linkages between each other by homologous recombination. By metaphase I, the chromatids of each replicated chromosome are cross-linked by cohesin complexes along their full length. Rec8, a meiosis-specific homolog of Scc1, is cleaved along chromosome arms but not around the centromere, allowing homologous chromosome pairs to segregate to daughter cells. Centromeric Rec8 is protected from cleavage by PP2A, recruited to centromeric regions by the PP2A regulator Mei-S332/Shugoshin (shown in purple). By metaphase II, the Mei-S332/Shugoshin-PP2A complex dissociates from chromosomes. Cohesins can now be cleaved during meiosis II, allowing sister chromatids to segregate. See F. Uhlmann, 2001, Curr. Opin. Cell Biol.13:754.
Page 914
The mechanisms that remove cohesins during meiosis are the same as during mitosis. Securin degradation releases separase, which then cleaves the cohesins holding the chromosome arms together, but leaves the cohesins around centromeres intact. This allows the recombined maternal and paternal chromosomes to separate, but each pair of chromatids remains associated at the centromere. During metaphase II, sister chromatids align on the metaphase II spindle, and separase is activated yet again, cleaving the residual cohesin around centromeres, facilitating anaphase II (see Figure 19-37b).