Sheet-Forming Type IV Collagen Is a Major Structural Component of the Basal Lamina
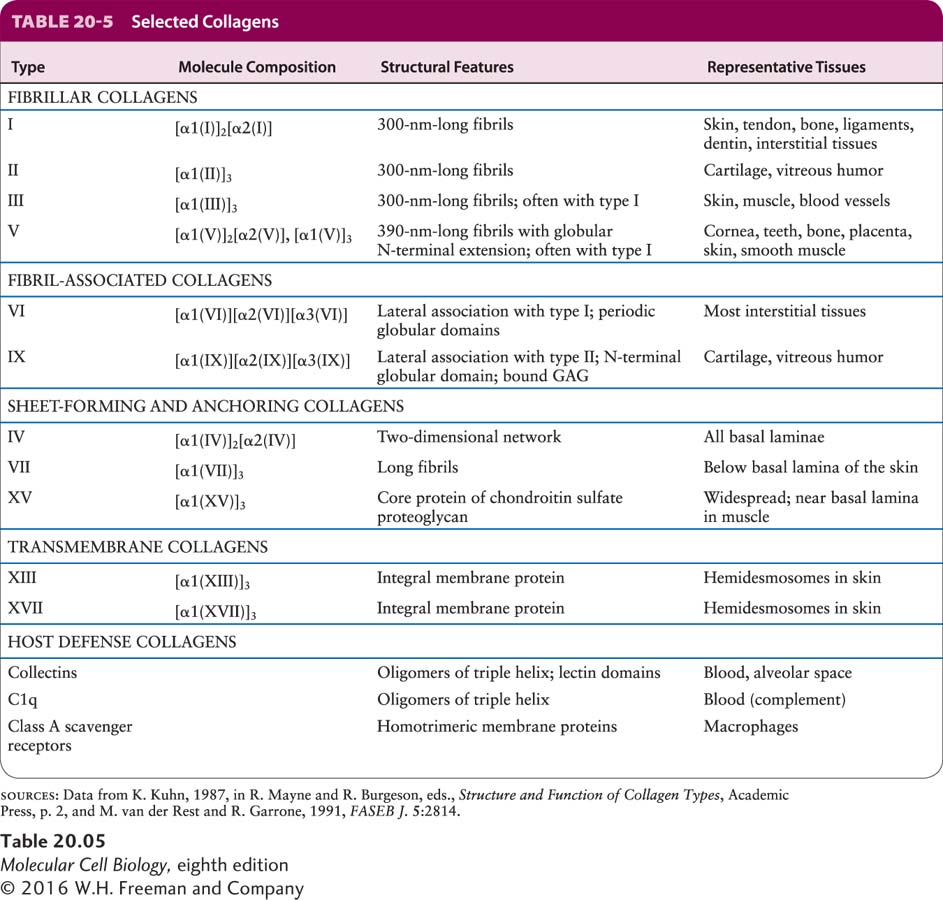
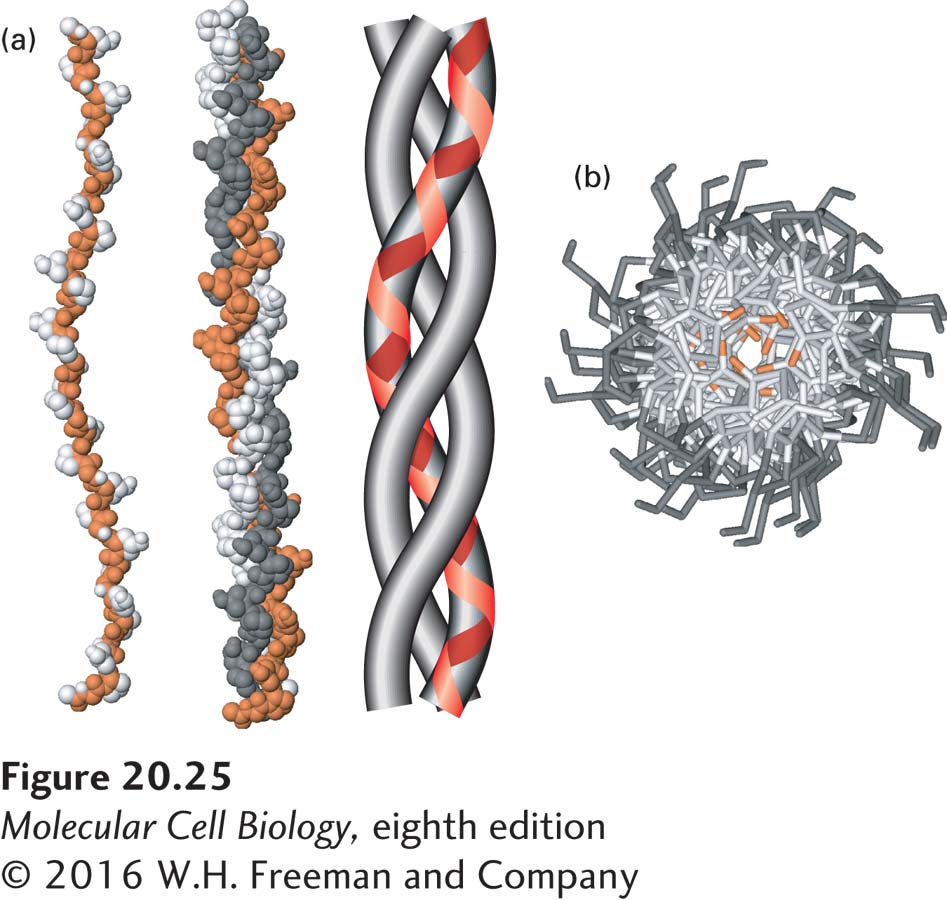
Type IV collagen is, together with laminin, a principal structural component of all basal laminae and can bind to adhesion receptors, including some integrins. Collagen IV is one of at least 28 types of collagen in humans that participate in the formation of distinct ECMs in various tissues (Table 20-5). There are also at least 20 additional collagen-like proteins (such as host defense collagens) in the human proteome. Although collagen isoforms differ in certain structural features and in their tissue distribution, all collagens are trimeric proteins made from three polypeptides, each encoded by one of at least 43 genes in humans, usually called collagen α chains. The three α chains in a collagen molecule can be identical (forming a homotrimer) or different (forming a heterotrimer). All or parts of the three-stranded collagen molecule can twist together into a special triple helix called a collagenous triple helix. When there is more than one triple-helical segment, these segments are joined by nonhelical regions of the protein, as we will see shortly for type IV collagen. Within a helical segment, each of the three α chains twists into a left-handed helix, and the three chains then wrap around one another to form a right-handed triple helix (Figure 20-25).
FIGURE 20-25The collagen triple helix. (a) Left: Side view of the crystal structure of a polypeptide fragment whose sequence is based on repeating sets of three amino acids, Gly-X-Y, characteristic of collagen α chains. Center: Each chain is twisted into a left-handed helix, and three chains wrap around one another to form a right-handed triple helix. The schematic model (right) clearly illustrates the triple-helical nature of the structure and shows the left-handed twist of the individual collagen α chains (red line). (b) View down the axis of the triple helix. The proton side chains of the glycine residues (orange) point into the very narrow space between the polypeptide chains in the center of the triple helix. In collagen mutations in which other amino acids replace glycine, the proton in glycine is replaced by larger groups that disrupt the packing of the chains and destabilize the triple-helical structure. Data from R. Z. Kramer et al., 2001, J. Mol. Biol.311:131, PDB ID 1bkv.
The collagen triple helix can form because of an unusual abundance of three amino acids in the α chains: glycine, proline, and a modified form of proline called hydroxyproline (see Figure 2-15). They make up the characteristic repeating sequence motif Gly-X-Y, where X and Y can be any amino acid but are often proline in position X and hydroxyproline in position Y, and less often lysine and hydroxylysine. Glycine is essential because its small side chain, a hydrogen atom, is the only one that can fit into the crowded center of the three-stranded helix (see Figure 20-25b). Hydrogen bonds help hold the three chains together. Although the rigid peptidyl-proline and peptidyl-hydroxyproline linkages are not compatible with formation of a classic single-stranded α helix, they stabilize the distinctive collagenous triple helix. The hydroxyl group in hydroxyproline in the Y position helps hold its ring in a conformation that stabilizes the three-stranded helix.
Page 949
There are several distinct cell-surface receptors for collagen IV and other types of collagen (other collagens are discussed in the next section). These cell-surface receptors include certain integrins, discoidin domain receptors 1 and 2 (which are tyrosine kinase receptors), glycoprotein VI (on platelets), leukocyte-associated Ig-like receptor-1, members of the mannose receptor family, and a modified form of the protein CD44. They can play critical roles in helping to assemble the ECM and in integrating cellular activity with the ECM.
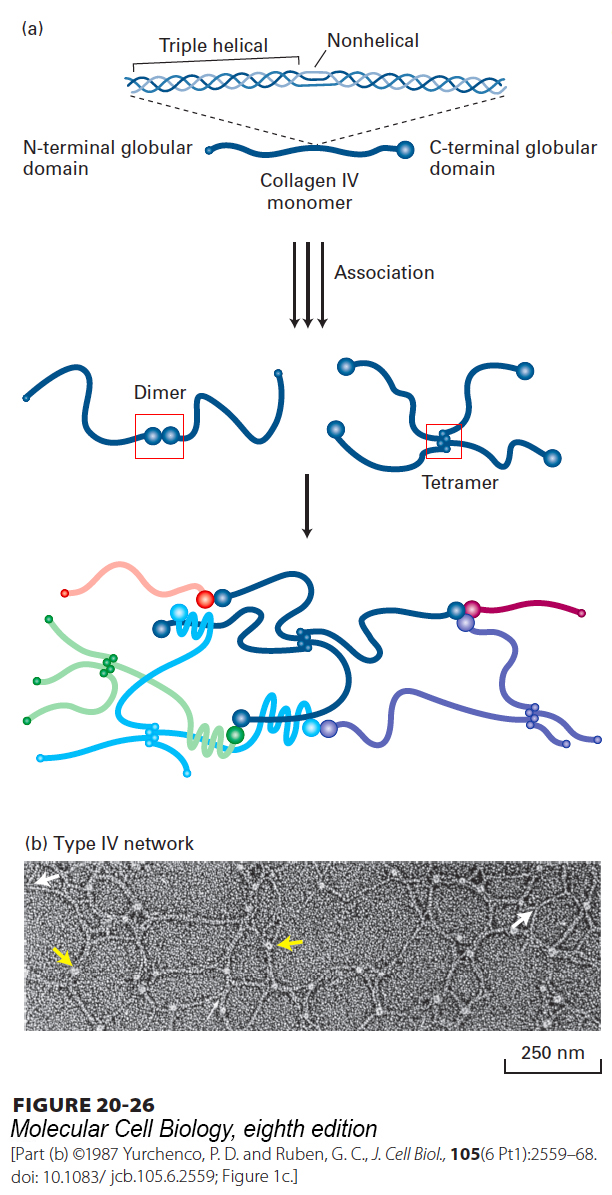
The unique properties of each collagen isoform are due mainly to differences in (1) the number and lengths of the collagenous triple-helical segments; (2) the segments that flank or interrupt the triple-helical segments and that fold into other kinds of three-dimensional structures; and (3) covalent modification of the α chains (e.g., hydroxylation, glycosylation, oxidation, cross-linking). For example, the chains in type IV collagen are designated IVα chains. Mammals express six homologous IVα chains, which assemble into three different heterotrimeric type IV collagens with distinct properties. All subtypes of type IV collagen, however, form a 400-nm-long triple helix (Figure 20-26) that is interrupted about 24 times by nonhelical segments and flanked by a large globular domain at the C-terminus of the chain and a smaller globular domain at the N-terminus. The nonhelical regions introduce flexibility into the molecule. Through both lateral associations and interactions entailing the globular N- and C-termini, type IV collagen molecules assemble into a branching, irregular two-dimensional fibrous network that forms a lattice on which, together with the laminin lattice, the basal lamina is built (see Figures 20-23 and 20-26).
FIGURE 20-26Structure and assembly of type IV collagen. (a) Schematic representation of type IV collagen. This 400-nm-long molecule has a small noncollagenous globular domain at the N-terminus and a large globular domain at the C-terminus. The collagenous triple helix is interrupted by nonhelical segments that introduce flexible kinks into the molecule. Lateral interactions between triple-helical segments, as well as head-to-head and tail-to-tail interactions between the globular domains, form dimers, tetramers, and higher-order complexes, yielding a sheet-like network. Multiple, unusual sulfilimine (–S=N–) or thioether bonds between hydroxylysine (or lysine) and methionine residues covalently cross-link some adjacent C-terminal domains and contribute to the stability of the network. See A. Boutaud, 2000, J. Biol. Chem.275:30716. (b) Electron micrograph of type IV collagen network formed in vitro. The lacy appearance results from the flexibility of the molecule, the side-to-side binding between triple-helical segments (white arrows), and the interactions between C-terminal globular domains (yellow arrows).
In the kidney, a double basal lamina called the glomerular basement membrane separates the epithelium that lines the urinary space from the endothelium that lines the surrounding blood-filled capillaries. Defects in this structure, which is responsible for ultrafiltration of the blood and initial urine formation, can lead to renal failure. For instance, mutations that alter the C-terminal globular domain of certain IVα chains are associated with progressive renal failure as well as sensorineural hearing loss and ocular abnormalities, a condition known as Alport’s syndrome. In Goodpasture’s syndrome, a relatively rare autoimmune disease, antibodies bind to the α3 chains of type IV collagen found in the glomerular basement membrane and lungs. This binding sets off an immune response that causes cellular damage, resulting in progressive renal failure and pulmonary hemorrhage.