Fibrillar Collagens Are the Major Fibrous Proteins in the ECM of Connective Tissues
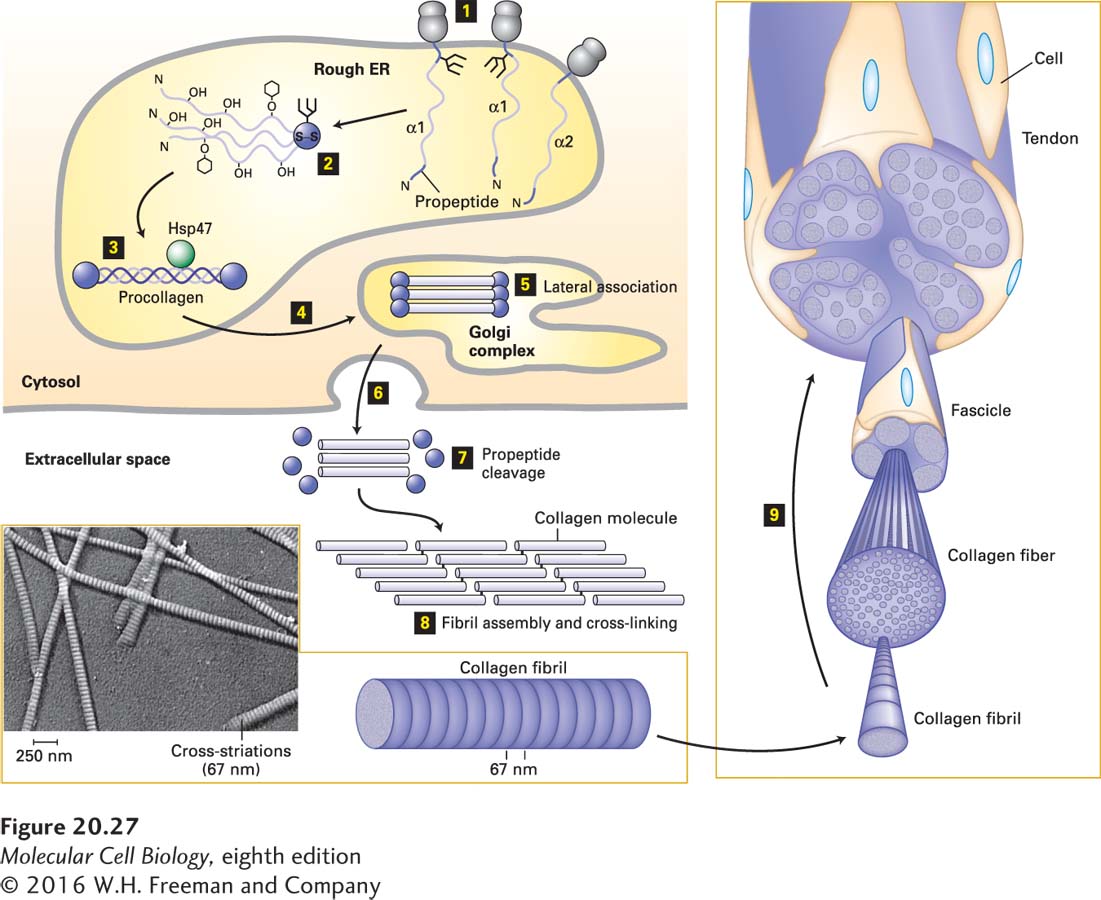
About 80–90 percent of the collagen in the body consists of fibrillar collagens (types I, II, and III), located primarily in connective tissues (see Table 20-5). Because of its abundance in tendon-rich tissue such as rat tail, type I collagen is easy to isolate and was the first collagen to be characterized. Its fundamental structural unit is a long (300-nm), thin (1.5-nm-diameter) triple helix (see Figure 20-25) consisting of two α1(I) chains and one α2(I) chain, each 1050 amino acids in length. The triple-stranded molecules pack tightly together and wrap around one another, forming microfibrils that associate into higher-order polymers called collagen fibrils, which in turn often aggregate into larger bundles called collagen fibers (Figure 20-27).
[Inset: Republished by permission of John Wiley & Sons, Inc., from Gross, J., “Evaluation of structural and chemical changes in connective tissue,” Ann. NY Acad. Sci., 1953, 56(4):674–83; permission conveyed through the Copyright Clearance Center, Inc.]
FIGURE 20-27Biosynthesis of fibrillar collagens. Step 1: Procollagen α chains are synthesized on ribosomes associated with the endoplasmic reticulum (rough ER), and in the ER, asparagine-linked oligosaccharides are added to the C-terminal propeptide. Step 2: Propeptides associate to form trimers and are covalently linked by disulfide bonds, and selected residues in the Gly-X-Y triplet repeats are covalently modified [certain prolines and lysines are hydroxylated, galactose or galactose-glucose (hexagons) are attached to some hydroxylysines, prolines are cis → trans isomerized]. Step 3: The modifications facilitate zipper-like formation and stabilization of triple helices, and binding by the chaperone protein Hsp47, which may stabilize the helices or prevent premature aggregation of the trimers, or both. Steps 4 and 5: The folded procollagens are transported to and through the Golgi complex, where some lateral association into small bundles takes place. The chains are then secreted (step 6), the N- and C-terminal propeptides are removed (step 7), and the trimers assemble into fibrils and are covalently cross-linked (step 8). The 67-nm staggering of the trimers gives the fibrils a striated appearance in electron micrographs (inset). Step 9: The fibrils can assemble into larger and larger bundles, some of which form the tendons that attach muscle to bone. See A. V. Persikov and B. Brodsky, 2002, Proc. Natl. Acad. Sci. USA99:1101–1103.
[Inset: Republished by permission of John Wiley & Sons, Inc., from Gross, J., “Evaluation of structural and chemical changes in connective tissue,” Ann. NY Acad. Sci., 1953, 56(4):674–83; permission conveyed through the Copyright Clearance Center, Inc.]
Classes of collagen that are less abundant, but nevertheless important, include fibril-associated collagens, which link the fibrillar collagens to one another or to other ECM components; sheet-forming and anchoring collagens, which form two-dimensional networks in basal laminae (type IV) and connect the basal lamina in skin to the underlying connective tissue (type VII); transmembrane collagens, which function as adhesion receptors; and host defense collagens, which help the body recognize and eliminate pathogens. Interestingly, several collagens (e.g., types IX, XVIII, and XV) are also proteoglycans with covalently attached GAGs (see Table 20-5).