Elastic Fibers Permit Many Tissues to Undergo Repeated Stretching and Recoiling
Elastic fibers are found in the ECM of a wide variety of tissues that are subject to mechanical strain or deformation, such as the lungs, which expand and contract during breathing (Figure 20-36a); the blood vessels, through which blood pulses due to the heartbeat, and the skin and many other tissues that stretch and contract. Elastic fibers permit the rubberlike reversible elastic stretching and recoiling of these tissues.
[Part (a) Biophoto Associates/Science Photo Library. Parts (b) and (c) Republished by permission of Elsevier, from Choi, J., et al., “Analysis of dermal elastic fibers in the absence of fibulin-5 reveals potential roles for fibulin-5 in elastic fiber assembly,” Matrix Biol., 2009, 28 (4):211–20; permission conveyed through the Copyright Clearance Center, Inc.]
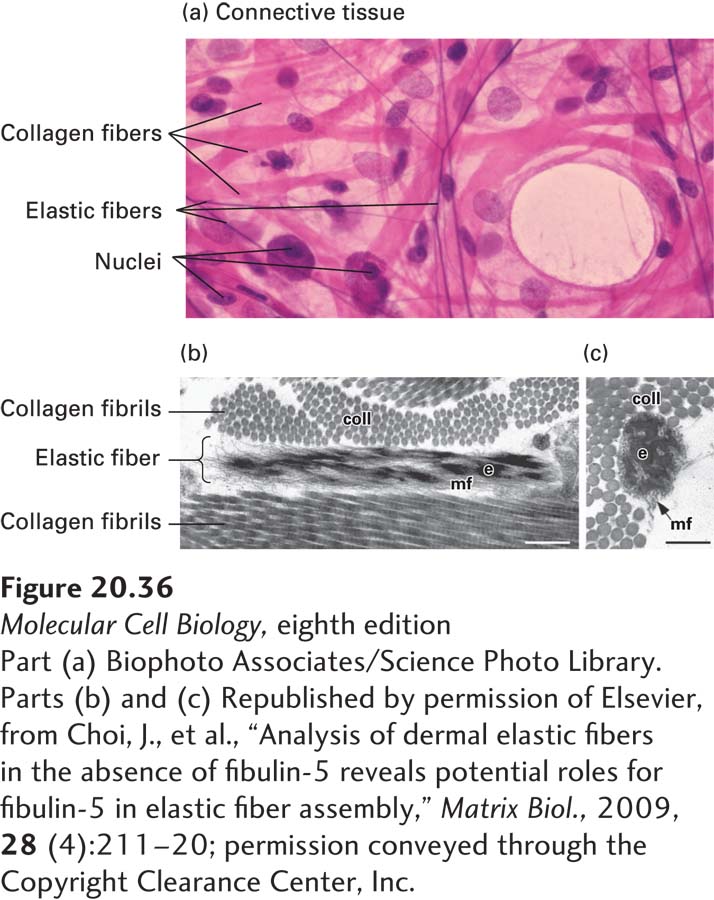
FIGURE 20-36Elastic and collagen fibers in connective tissue. (a) Light-microscopic image of loose connective tissue from the lung. Elastic fibers are the thin fibers that are stained purple, collagen fibers (bundles of collagen fibrils) are stained pink, and the nuclei of cells are stained purple. (b) Longitudinal and (c) cross-sectional electron microscopic images of elastic fibers and collagen fibrils (coll) in the skin of a mouse. The elastic fibers have a solid core of elastin (e) integrated into and surrounded by a bundle of microfibrils (mf). Scale bars, 0.25 μm.
[Part (a) Biophoto Associates/Science Photo Library. Parts (b) and (c) Republished by permission of Elsevier, from Choi, J., et al., “Analysis of dermal elastic fibers in the absence of fibulin-5 reveals potential roles for fibulin-5 in elastic fiber assembly,” Matrix Biol., 2009, 28 (4):211–20; permission conveyed through the Copyright Clearance Center, Inc.]
Page 960
The major component of an elastic fiber, which can be several hundred to several thousand nanometers in diameter, is an insoluble, amorphous core composed of the protein elastin. Elastin consists of aggregates of monomeric tropoelastin molecules that are covalently cross-linked via a lysyl oxidase–mediated process similar to that seen in collagen. Repetitive proline- and glycine-enriched hydrophobic sequence motifs contribute to the ability of tropoelastins to self-associate, extend under stress, and recoil efficiently after stretching. The elastin core is surrounded by a collection of 10–12-nm-diameter microfibrils made up of the proteins fibrillin, fibulin, and associated proteins such as LTBPs (Figure 20-36b). The microfibrils serve as scaffolds for the assembly of the elastic fiber’s core. Elastin-free microfibrils are found in the eye, where they transmit muscular force to reshape the lens for focusing and may provide structural support to the cornea.
Similar to other components of the ECM, microfibrils participate in cell signaling. In the secretory pathway, LTBPs bind the inactive form of transforming growth factor β (TGF-β; see Chapter 16) prior to their co-secretion and incorporation into microfibrils (indeed, LTBP is an acronym for latent TGF-β binding protein). Biomechanical stress mediated by cell-surface integrins binding to and pulling on the LTBP/TGF-β complex or proteolytic cleavage are thought to be the direct cause of active TGF-β release from the ECM and subsequent signaling (see Figure 16-3).
A variety of diseases, many involving skeletal and cardiovascular abnormalities, are consequences of mutations in the genes encoding the structural proteins of elastic fibers or the proteins that contribute to their proper assembly. For example, mutations in the fibrillin-1 gene cause Marfan syndrome, whose varied symptoms can include bone overgrowth, loose joints, abnormally long extremities and face, and cardiovascular defects due to weakness in the walls of the aorta and other blood vessels. There has been considerable speculation that President Abraham Lincoln’s unusually tall, elongated body may have been a consequence of Marfan syndrome.
In mammals, most tropoelastin synthesis occurs immediately before and after birth during the late fetal and neonatal periods. Thus most of the body’s elastin must be very durable, lasting an entire lifetime. The extraordinary stability of elastin has been measured in a variety of ways. Pulse-chase experiments (see Chapter 3) using radioactive amino acid administration can be used to measure the life span of elastin in animals. In humans, two other methods employed to study the longevity of elastin have revealed that the mean lifetime of an elastin molecule in human lungs is about 70 years! The first method takes advantage of a naturally occurring phenomenon: the slow, natural rate of conversion of L-aspartic acid—incorporated into proteins during their synthesis—to D-aspartic acid. Thus the age of a long-lived protein can be estimated using chemical analysis to determine the fraction of its L-aspartic acid that has been converted over time to the D isomer, together with knowledge of the age of the tissue from which it was isolated. The second method is a variation on the classic pulse-chase experiments used in the laboratory. As a consequence of nuclear weapons testing in the 1950s and 1960s, 14C was introduced into the atmosphere and hence the food chain. This environmental 14C has been used as the radioactive “pulse” in what is essentially a pulse-chase experiment to determine the stability of proteins of interest.