Integrins Mediate Adhesion and Relay Signals Between Cells and Their Three-Dimensional Environment
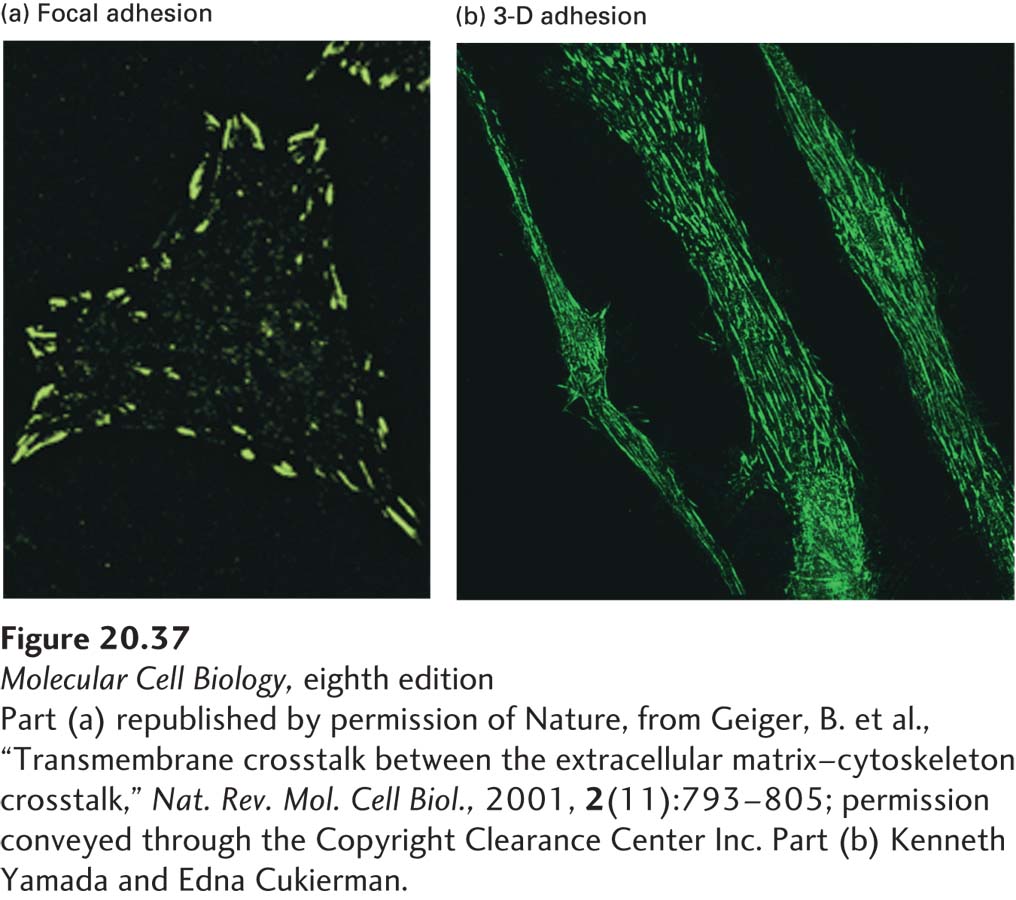
As already discussed, integrins connect epithelial cells to the basal lamina and, through adapter proteins, to intermediate filaments of the cytoskeleton (see Figure 20-1). That is, integrins form a bridge between the ECM and the cytoskeleton; they do the same in nonepithelial cells. In epithelial and nonepithelial cells, integrins in the plasma membrane are also clustered with other molecules in various focal contacts (focal adhesions) and focal contact–like adhesive structures called focal complexes, 3-D adhesions, and fibrillar adhesions, as well as in circular adhesions called podosomes. These structures are multiprotein complexes that mediate (1) cell adhesion to the ECM—for example, via integrin binding to fibronectin (see Figure 20-35) or laminin, (2) integrin association with the actin cytoskeleton, (3) adhesion-dependent outside-in and inside-out signaling (see Figure 20-8), and (4) mechanosensory coupling between cells and their environments. These complexes are readily observed by fluorescence microscopy with the use of antibodies that recognize integrins or other molecules clustered with them (Figure 20-37).
[Part (a) republished by permission of Nature, from Geiger, B. et al., “Transmembrane crosstalk between the extracellular matrix–cytoskeleton crosstalk,” Nat. Rev. Mol. Cell Biol., 2001, 2(11):793–805; permission conveyed through the Copyright Clearance Center Inc. Part (b) Kenneth Yamada and Edna Cukierman.]
EXPERIMENTAL FIGURE 20-37Integrins cluster into adhesive structures with various morphologies in nonepithelial cells. Immunofluorescence methods were used to detect integrin-containing adhesive structures (green) on cultured cells. Shown here are (a) focal adhesions and (b) 3-D adhesions on the surfaces of human fibroblasts. Cells were grown (a) directly on the flat surface of a culture dish or (b) on a three-dimensional matrix of ECM components. The shape, distribution, and composition of the integrin-based adhesions formed by cells vary depending on the cells’ environment.
[Part (a) republished by permission of Nature, from Geiger, B. et al., “Transmembrane crosstalk between the extracellular matrix–cytoskeleton crosstalk,” Nat. Rev. Mol. Cell Biol., 2001, 2(11):793–805; permission conveyed through the Copyright Clearance Center Inc. Part (b) Kenneth Yamada and Edna Cukierman.]
Integrin-containing adhesive structures are dynamic due to ongoing import, export, or covalent modification of their components, and each contains dozens of intracellular adapter and associated proteins. The hundreds of such proteins identified to date have the potential to engage in many hundreds of distinct protein-protein interactions that may be subject to regulation. For example, binding sites generated by phosphorylation of integrin and its associated proteins, as well as by generation of phosphorylated derivatives of phosphatidylinositol in the adjacent membrane, recruit additional proteins into, and can also cause release of some proteins from, these multiprotein complexes. A tightly controlled choreography of internal signals, contributions of other signaling pathways such as those involving receptor tyrosine kinases (see Figure 20-8), and external signals (such as the composition and rigidity of the ECM) regulates these complexes. Together, they help define the precise composition and activity of the integrin multiprotein complex and the consequent influence that it has on cellular structure and activity (outside-in effect) as well as the influence of the cellular actin cytoskeleton on the ECM (inside-out effect).
Page 962
Although found in many nonepithelial cells, integrin-containing adhesive structures have been studied most frequently in fibroblasts grown in cell culture on flat glass or plastic surfaces called substrata. These conditions only poorly approximate the three-dimensional ECM environment that normally surrounds such cells in vivo. When fibroblasts are cultured in three-dimensional ECMs derived from cells or tissues, they form adhesions to the three-dimensional ECM substratum, called 3-D adhesions. These structures differ somewhat in composition, shape, distribution, and activity from the focal or fibrillar adhesions seen in cells growing on the flat substrata typically used in cell culture experiments (see Figure 20-37). Cultured fibroblasts with these “more natural” anchoring junctions display greater adhesion and mobility, increased rates of cell proliferation, and spindle-shaped morphologies more like those of fibroblasts in tissues than do cells cultured on hard, flat surfaces. These and other observations indicate that the topological, compositional, and mechanical properties of the ECM all play a role in controlling the shape and activity of a cell. Tissue-specific differences in these ECM characteristics probably contribute to the tissue-specific properties of cells.
The importance of the three-dimensional environment of cells has been highlighted by cell culture studies of the morphogenesis, functioning, and stability of specialized milk-producing mammary epithelial cells and their cancerous transformed counterparts. For example, the three-dimensional ECM-dependent outside-in signaling mediated by integrins influences the epidermal growth factor–tyrosine kinase receptor signaling system, and vice versa. The three-dimensional ECM also permits the mammary epithelial cells to generate in vivo–like circular epithelial structures, called acini, which secrete the major protein constituents of milk. The use of such three-dimensional ECM cell culture systems permits more realistic comparisons of the responses of normal and cancer cells to potential chemotherapeutic agents. Analogous systems employing both natural and synthetic three-dimensional ECMs are being developed to provide more in vivo–like conditions to study other complex tissues and organs, such as the liver.