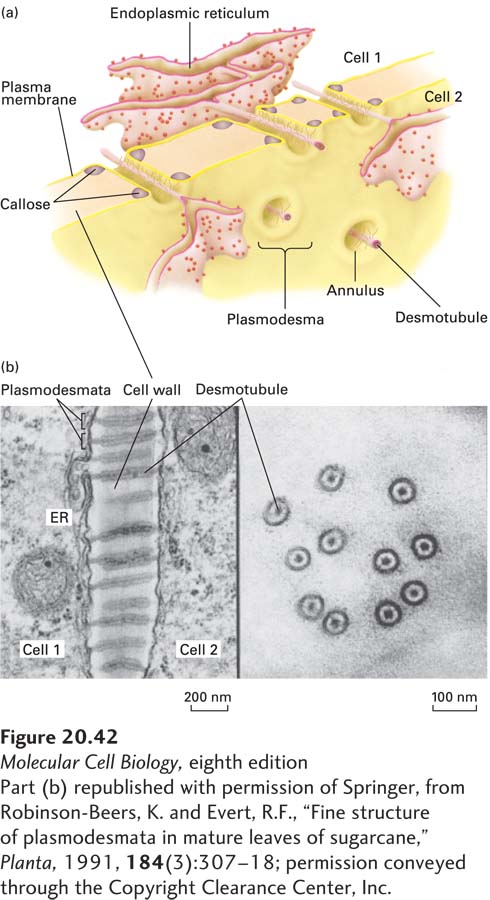
Plasmodesmata Directly Connect the Cytosols of Adjacent Cells
The presence of a cell wall separating cells in plants imposes barriers to cell-cell communication—and thus cell differentiation—not faced by animals. One distinctive mechanism used by plant cells to communicate directly is specialized cell junctions called plasmodesmata, which extend through the cell wall (Figure 20-42). Like gap junctions, plasmodesmata are channels that connect the cytosol of a cell with that of an adjacent cell. The diameter of the channel is about 30–60 nm, and its length can vary, but may be greater than 1 µm. The density of plasmodesmata varies depending on the plant and cell type, and even the smallest meristematic cells have more than a thousand connections with their neighbors. An adapter protein called NET1A is thought to link the plasmodesmata to the actin cytoskeleton. Although a variety of proteins and polysaccharides that are physically or functionally associated with plasmodesmata have been identified, the key structural protein components of plasmodesmata and the detailed mechanisms underlying their biogenesis remain to be identified.
[Part (b) republished with permission of Springer, from Robinson-Beers, K. and Evert, R.F., “Fine structure of plasmodesmata in mature leaves of sugarcane,” Planta, 1991, 184(3):307–18; permission conveyed through the Copyright Clearance Center, Inc.]
FIGURE 20-42Plasmodesmata. (a) Schematic model of plasmodesmata, showing the desmotubule, an extension of the endoplasmic reticulum (ER), and the annulus, a plasma-membrane-lined channel filled with cytosol that interconnects the cytosols of adjacent cells. The regulated deposition of a glucose polymer called callose (cyan) in the extracellular spaces in the cell wall adjacent to the entrances of the channels has the potential to block intercellular transport through the plasmodesmata, apparently by forcing the closing of the channels by narrowing the annulus. (b) Electron micrographs of thin sections of a sugarcane leaf (brackets indicate individual plasmodesmata). Left: Longitudinal view, showing ER and desmotubule running through each annulus. Right: Perpendicular cross-sectional views of plasmodesmata, in some of which spoke structures connecting the plasma membrane to the desmotubule can be seen.
[Part (b) republished with permission of Springer, from Robinson-Beers, K. and Evert, R.F., “Fine structure of plasmodesmata in mature leaves of sugarcane,” Planta, 1991, 184(3):307–18; permission conveyed through the Copyright Clearance Center, Inc.]
Molecules smaller than about 1000 Da, including a variety of metabolic and signaling compounds (ions, sugars, amino acids), can generally diffuse through plasmodesmata. However, the size of the channel through which molecules pass is highly regulated. In some circumstances, the channel is clamped shut; in others, it is dilated sufficiently to permit the passage of molecules larger than 10,000 Da. The deposition and breakdown of a glucose polymer called callose in the extracellular spaces adjacent to the entrances of the channels (see Figure 20-42a) is thought to regulate the closing and opening of the channels, respectively. Among the factors that affect the permeability of plasmodesmata is the cytosolic Ca2+ concentration: an increase in cytosolic Ca2+ reversibly inhibits movement of molecules through these structures.
Page 970
Although plasmodesmata and gap junctions resemble each other functionally with respect to forming channels for small-molecule diffusion, their structures differ dramatically in two significant ways (see Figure 20-42). In plasmodesmata, the plasma membranes of the adjacent plant cells merge to form a continuous channel, called the annulus, whereas the plasma membranes of animal cells at a gap junction are not continuous with each other. There are simple plasmodesmata (with a single pore, like those in Figure 20-42) and complex plasmodesmata that branch into multiple channels. In addition, plasmodesmata exhibit many additional complex structural and functional characteristics. For example, they contain within the channel an extension of the endoplasmic reticulum, called a desmotubule, that passes through the annulus. They also have a variety of specialized proteins at the entrance of the channel and running throughout the length of the channel, including cytoskeletal, motor, and docking proteins that regulate the sizes and types of molecules that can pass through the channel. Many types of molecules spread from cell to cell through plasmodesmata, including some transcription factors, nucleic acid/protein complexes, metabolic products, and plant viruses. It appears that some of these require special chaperones to facilitate transport. Specialized kinases may also phosphorylate plasmodesmal components to regulate their activities (e.g., opening of the channels). Soluble molecules pass through the cytosolic annulus, about 3–4 nm in diameter, that lies between the plasma membrane and desmotubule, whereas membrane-bound molecules or certain proteins within the ER lumen can pass from cell to cell via the desmotubule. Plasmodesmata appear to play an especially important role in protection from pathogens and in regulating the development of plant cells and tissues, as is suggested by their ability to mediate intracellular movement of transcription factors and ribonuclear protein complexes.