Only a Few Adhesion Molecules Have Been Identified in Plants
[Republished with permission of Springer, from Guang Yuh, J., et al., “Adhesion of lily pollen tubes on an artificial matrix,” Sex. Plant Reprod., 1997, 10:3, pp. 173–180.]
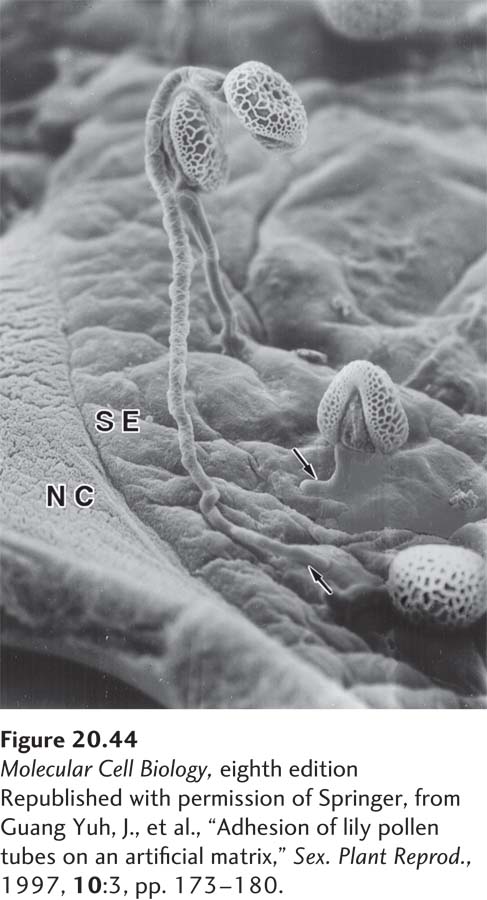
EXPERIMENTAL FIGURE 20-44An in vitro assay was used to identify molecules required for adherence of pollen tubes to the stylar ECM. In this assay, ECM collected from lily styles (SE) or an artificial ECM was dried on nitrocellulose membranes (NC). Pollen tubes containing sperm were then added, and their binding to the dried ECM was assessed. In this scanning electron micrograph, the tips of pollen tubes (arrows) can be seen binding to dried stylar ECM. This type of assay has shown that pollen adherence depends on stigma/stylar cysteine-rich adhesin (SCA) and a pectin that binds to SCA.
[Republished with permission of Springer, from Guang Yuh, J., et al., “Adhesion of lily pollen tubes on an artificial matrix,” Sex. Plant Reprod., 1997, 10:3, pp. 173–180.]
Systematic analyses of the Arabidopsis genome and biochemical analyses of other plant species have provided no evidence for the existence of plant homologs of most animal CAMs, adhesion receptors, and ECM components. This finding is not surprising, given the dramatically different nature of cell-cell and cell-ECM interactions in animals and plants.
Among the adhesive proteins apparently unique to plants are five wall-associated kinases (WAKs) and WAK-like proteins expressed in the plasma membrane of Arabidopsis cells. These transmembrane proteins have a cytoplasmic serine/threonine kinase domain, and their extracellular regions contain multiple epidermal growth factor (EGF) repeats, frequently found in animal cell-surface receptors. Some WAKs have an extracellular pectin-binding domain that can recognize and bind full-length pectin and pectin degradation fragments. Such binding has been proposed to help cells monitor and respond to the status of the cell wall during normal growth and in the context of cell-wall damage (wounding) or infection by pathogens. Thus some WAKS in plant cells appear to be analogous to adhesion receptors in animal cells, binding and sensing the ECM and mediating outside-in signaling.
The results of in vitro binding assays, combined with in vivo studies and analyses of plant mutants, have identified several macromolecules in the ECM that are important for adhesion. For example, normal adhesion of pollen, which contains sperm cells, to the stigma or style in the female reproductive organ of the Easter lily requires a cysteine-rich protein called stigma/stylar cysteine-rich adhesin (SCA) and a specialized pectin that can bind to SCA (Figure 20-44). A small, probably ECM-embedded, 10-kDa protein called chymocyanin works in conjunction with SCA to help direct the movement of the sperm-containing pollen tube (chemotaxis) to the ovary.
Disruption of the gene encoding glucuronyltransferase 1, a key enzyme in pectin biosynthesis, has provided a striking illustration of the importance of pectins in intercellular adhesion in plant meristems. Normally, specialized pectin molecules help hold the cells in meristems tightly together. When grown in culture as a cluster of relatively undifferentiated cells, called a callus, normal meristematic cells adhere tightly and can differentiate into chlorophyll-producing cells, giving the callus a green color. Eventually the callus will generate shoots. In contrast, mutant cells with an inactivated glucuronyltransferase 1 gene are large, associate loosely with one another, and do not differentiate normally, forming a yellow callus. The introduction of a normal glucuronyltransferase 1 gene into the mutant cells restores their ability to adhere and differentiate normally.
Page 972
The paucity of plant adhesion molecules identified to date, in contrast to the many well-defined animal adhesion molecules, may be due to the technical difficulties of working with the ECM/cell wall of plants. Adhesive interactions are likely to play different roles in plant and animal biology, at least in part because of the differences in development and physiology between plants and animals.