Intestinal Stem Cells Continuously Generate All the Cells of the Intestinal Epithelium
The epithelium lining the small intestine is a single cell thick (see Figure 20-11) and is composed of four types of differentiated cells. The most abundant epithelial cells, the absorptive enterocytes, transport nutrients essential for survival from the intestinal lumen into the body (see Figure 11-30). The intestinal epithelium is the most rapidly self-renewing tissue in adult mammals, turning over every 5 days; in humans, up to 300,000,000 intestinal epithelial cells, weighing a total of about 1 gram, are lost every day.
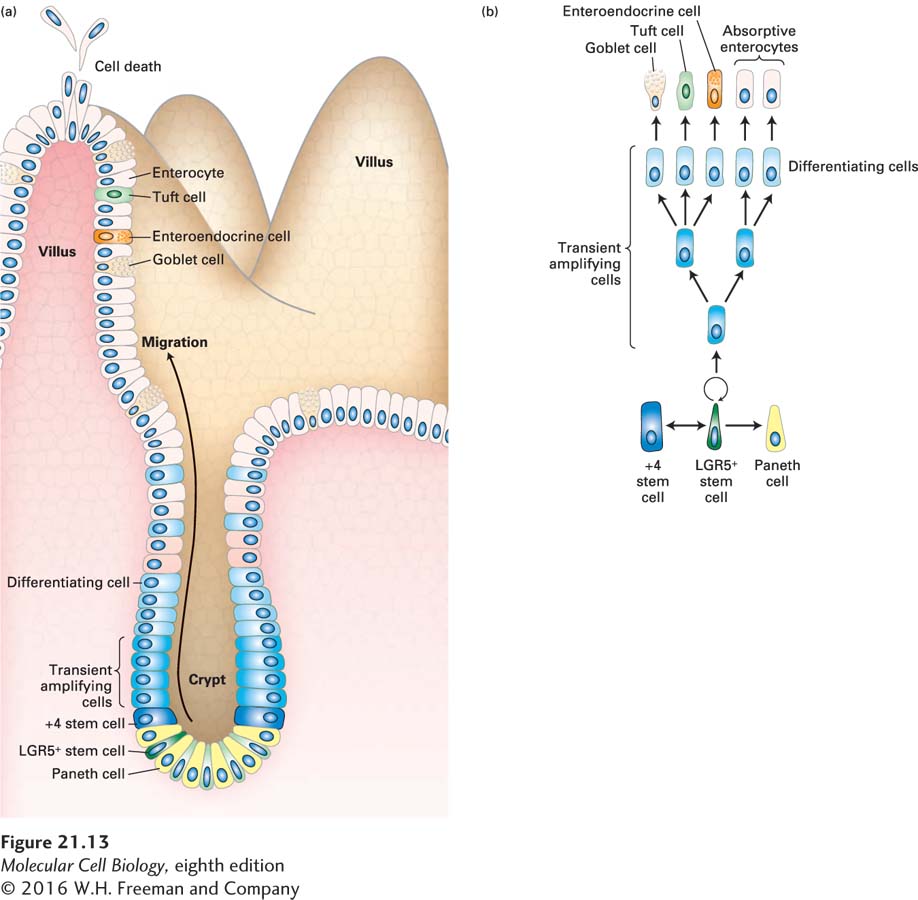
The cells of the intestinal epithelium are continuously regenerated from a stem-cell population located deep in the intestinal wall in pits called crypts (Figure 21-13). Pulse-chase experiments using radiolabeled thymidine have shown that intestinal stem cells produce precursor cells that divide rapidly and then differentiate as they ascend the sides of crypts to form the surface layer of the fingerlike gut projections called villi, across which intestinal absorption occurs. The time from cell birth in the crypts to the loss of dead cells at the tips of the villi is only about 3 to 5 days (Figure 21-14). The production of new cells is precisely controlled: too little division would eliminate villi and lead to breakdown of the intestinal surface; too much division would create an excessively large epithelium and might also be a step toward cancer.
FIGURE 21-13Intestinal stem cells and their niche. (a) Schematic drawing of an intestinal crypt and villus, showing the Lgr5-expressing (Lgr5+) intestinal stem cells (dark green), their mitotic progeny, the transient amplifying cells (intermediate blue), the terminal differentiating cells (light blue), and the several types of differentiated cells in the villus. The base of the crypt is the location of Paneth cells (yellow), which provide a major part of the stem-cell niche and also secrete antimicrobial defense proteins. The +4 “reserve” stem cells (which occupy the fourth position from the crypt base, dark blue) can restore the Lgr5+ stem-cell compartment following injury and can also be generated from these stem cells. (b) Lineages of cells in the small intestine. Epithelial turnover occurs every 3–5 days. New Paneth cells are supplied from the transient amplifying cells every 3–6 weeks. See N. Barker, 2014, Nat. Rev. Mol. Cell Biol.15:19.
[Republished with permission of John Wiley & Sons, Inc., from Kaur, P. and Potten, C. S., “Cell migration velocities in the crypts of the small intestine after cytotoxic insult are not dependent on mitotic activity,” Cell Tissue Kinet., 1986, 6:601–610; permission conveyed through Copyright Clearance Center, Inc.]
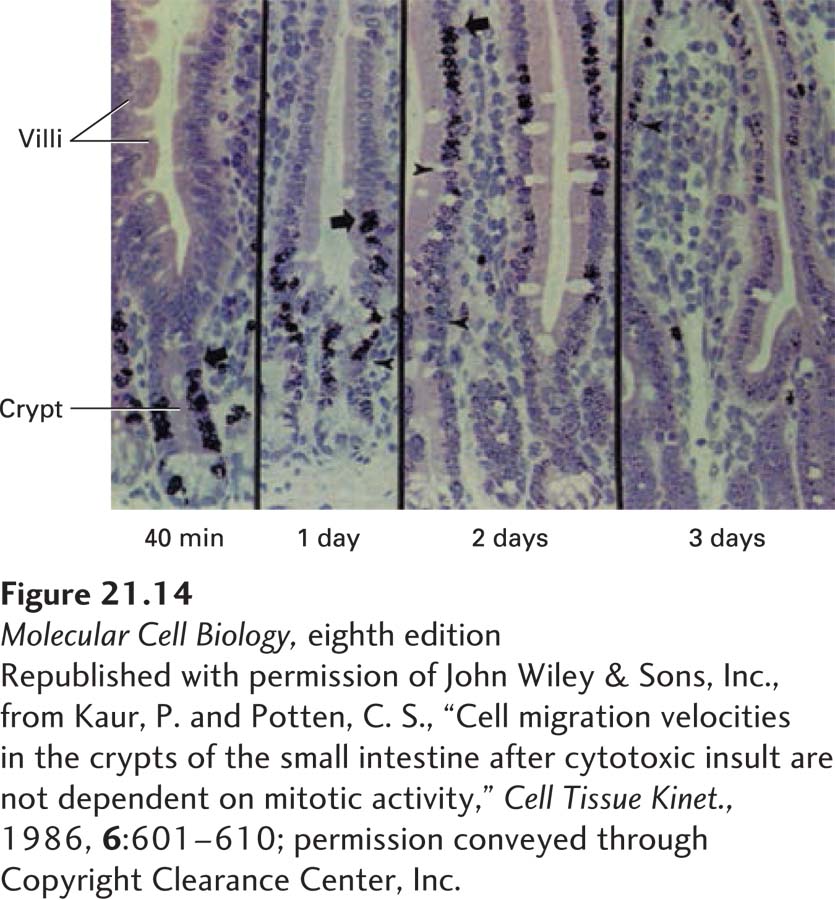
EXPERIMENTAL FIGURE 21-14Regeneration of the intestinal epithelium from stem cells can be demonstrated in pulse-chase experiments. Results from a pulse-chase experiment in which radioactively labeled thymidine (the pulse) was added to a culture of intestinal epithelial tissue. Dividing cells incorporated the labeled thymidine into their newly synthesized DNA. The labeled thymidine was washed away and replaced with unlabeled thymidine (the chase) after a brief period; cells that divided after the chase did not become labeled. These micrographs show that 40 minutes after labeling, all of the label is in cells near the base of the crypt. At later times, the labeled cells are seen progressively farther away from their point of birth in the crypt. Cells at the top are shed. This process ensures constant replenishment of the gut epithelium with new cells.
[Republished with permission of John Wiley & Sons, Inc., from Kaur, P. and Potten, C. S., “Cell migration velocities in the crypts of the small intestine after cytotoxic insult are not dependent on mitotic activity,” Cell Tissue Kinet., 1986, 6:601–610; permission conveyed through Copyright Clearance Center, Inc.]
Page 992
Experiments such as the one depicted in Figure 21-14 suggested that the intestinal stem cells were located somewhere near the bottom of the crypts, near differentiated intestinal cells called Paneth cells. But these putative stem cells had no particular morphological characteristics that revealed their remarkable abilities; which cells were the actual intestinal stem cells and which were the supportive cells that form the niche?
Prior genetic experiments had shown that Wnt signals are essential for intestinal stem-cell maintenance. As evidence for the importance of these signals, overproduction of active β-catenin (normally activated by the Wnt signaling pathway; see Figure 16-30) in intestinal cells leads to excess proliferation of the intestinal epithelium. Conversely, blocking the function of β-catenin by mutating or inhibiting the Wnt-activated TCF transcription factor abolishes the stem cells in the intestine, leading to intestinal degeneration and eventual death. Thus Wnt signaling plays a critical role in the intestinal stem-cell niche, as it does in the skin, blood, and other organs. Indeed, mutations that inappropriately activate the Wnt signaling pathway are a major contributor to the progression of colon cancer, as we will see in Chapter 24.
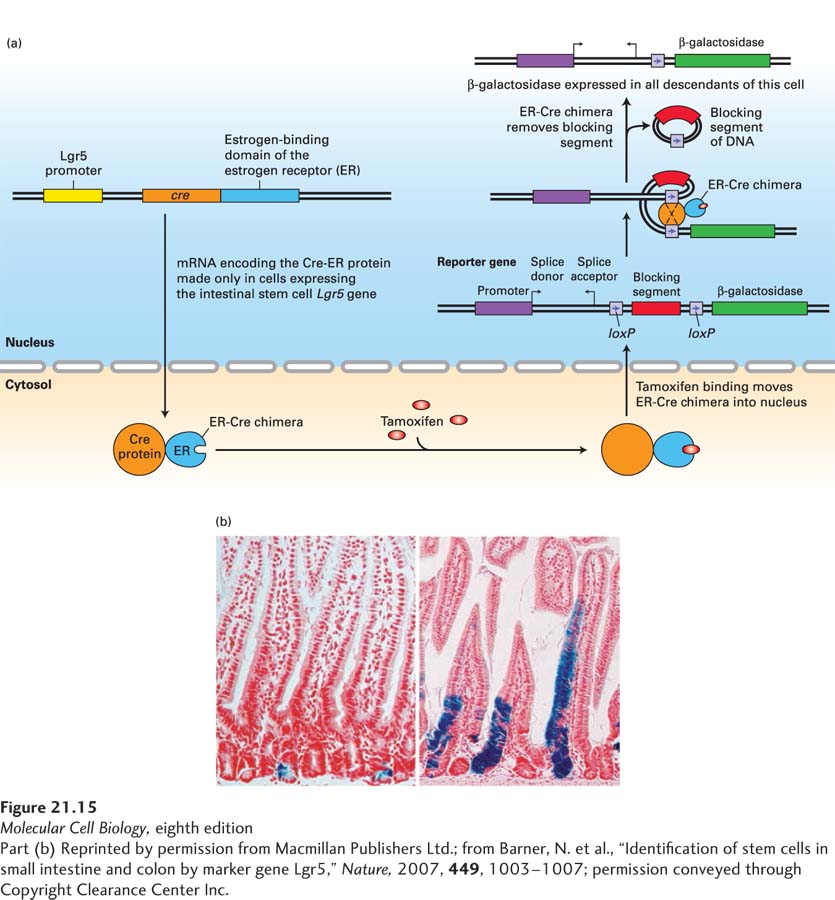
By analyzing a panel of genes whose expression in the intestine was induced by Wnt signaling, investigators zeroed in on Lgr5, a gene encoding a G protein–coupled receptor, because it was expressed only in a small set of cells at the very base of the crypts. Lgr5 binds a class of secreted hormones termed R-spondins and activates intracellular signaling pathways that potentiate Wnt signaling. Lineage-tracing studies showed that the descendants of these Lgr5-expressing cells indeed gave rise to all of the differentiated intestinal epithelial cells (Figure 21-15). These studies made use of genetically altered mice in which a version of the Cre recombination protein (see Figure 6-39), an estrogen receptor (ER)–Cre recombinase chimera, was placed under the control of the Lgr5 promoter; thus the ER-Cre recombinase chimera was produced only in the few putative Lgr5-expressing stem cells at the bottom of the crypts. The version of Cre recombinase used in the study had been altered so that it resides inactive in the cytosol and is transferred into the nucleus only after addition of an estrogen analog (Figure 21-15a). There the Cre excises a segment of DNA, activating expression of a β-galactosidase reporter gene. Importantly, all of the descendants of these cells will also express β-galactosidase. Immediately after addition of the estrogen analog, the only cells expressing β-galactosidase are the stem cells in the crypts. But after a few days, all of the descendant epithelial cells also expressed β-galactosidase (Figure 21-15b), showing that Lgr5 expression is indeed a marker of the intestinal stem cells.
[Part (b) Reprinted by permission from Macmillan Publishers Ltd.; from Barner, N. et al., “Identification of stem cells in small intestine and colon by marker gene Lgr5,” Nature, 2007, 449, 1003–1007; permission conveyed through Copyright Clearance Center Inc.]
EXPERIMENTAL FIGURE 21-15Lineage-tracing studies show that the Lgr5-expressing cells at the bases of crypts are the intestinal stem cells. (a) Outline of the experiment. Using genetically altered ES cells (see Figure 6-37), investigators generated one strain of mice in which a version of the gene encoding Cre recombinase (see Figure 6-39) was placed under the control of the Lgr5 promoter, and thus Cre recombinase was produced only in cells, such as intestinal stem cells, that express the Lgr5 gene. This version of Cre recombinase contained an additional domain from the estrogen receptor (ER) that binds the estrogen analog tamoxifen; like the estrogen receptor and other nuclear receptors (see Figure 9-45), the ER-Cre chimera is retained in the cytosol unless tamoxifen is added. In the presence of tamoxifen, ER-Cre moves into the nucleus, where it can interact with loxP sites in the chromosomal DNA. A second reporter strain of mice contained a bacterial β-galactosidase reporter gene that was preceded by two loxP sites. The blocking segment of DNA in between these loxP sites prevented expression of the β-galactosidase gene, and the β-galactosidase gene could be expressed only in cells where an active Cre recombinase had removed the sequence in between the two loxP sites. The two strains of mice were mated, and offspring containing both marker transgenes were identified. In these mice, β-galactosidase was expressed only in cells in which the Lgr5-controlled ER-Cre gene was expressed, and only after the estrogen analog tamoxifen was given to the mice. Thus only Lgr5-expressing cells—and all of their descendants—would express the β-galactosidase gene. (b) Results of the experiment. One day after tamoxifen was given to these mice, the only cells expressing β-galactosidase (indicated by the blue histochemical stain) were the Lgr5-expressing intestinal stem cells at the bases of the crypts (left). Five days after tamoxifen administration, additional blue cells—the epithelial descendants of the intestinal stem cells—were seen migrating up the sides of the villi. Some blue stem cells remained at the bottom of the crypt.
[Part (b) Reprinted by permission from Macmillan Publishers Ltd.; from Barner, N. et al., “Identification of stem cells in small intestine and colon by marker gene Lgr5,” Nature, 2007, 449, 1003–1007; permission conveyed through Copyright Clearance Center Inc.]
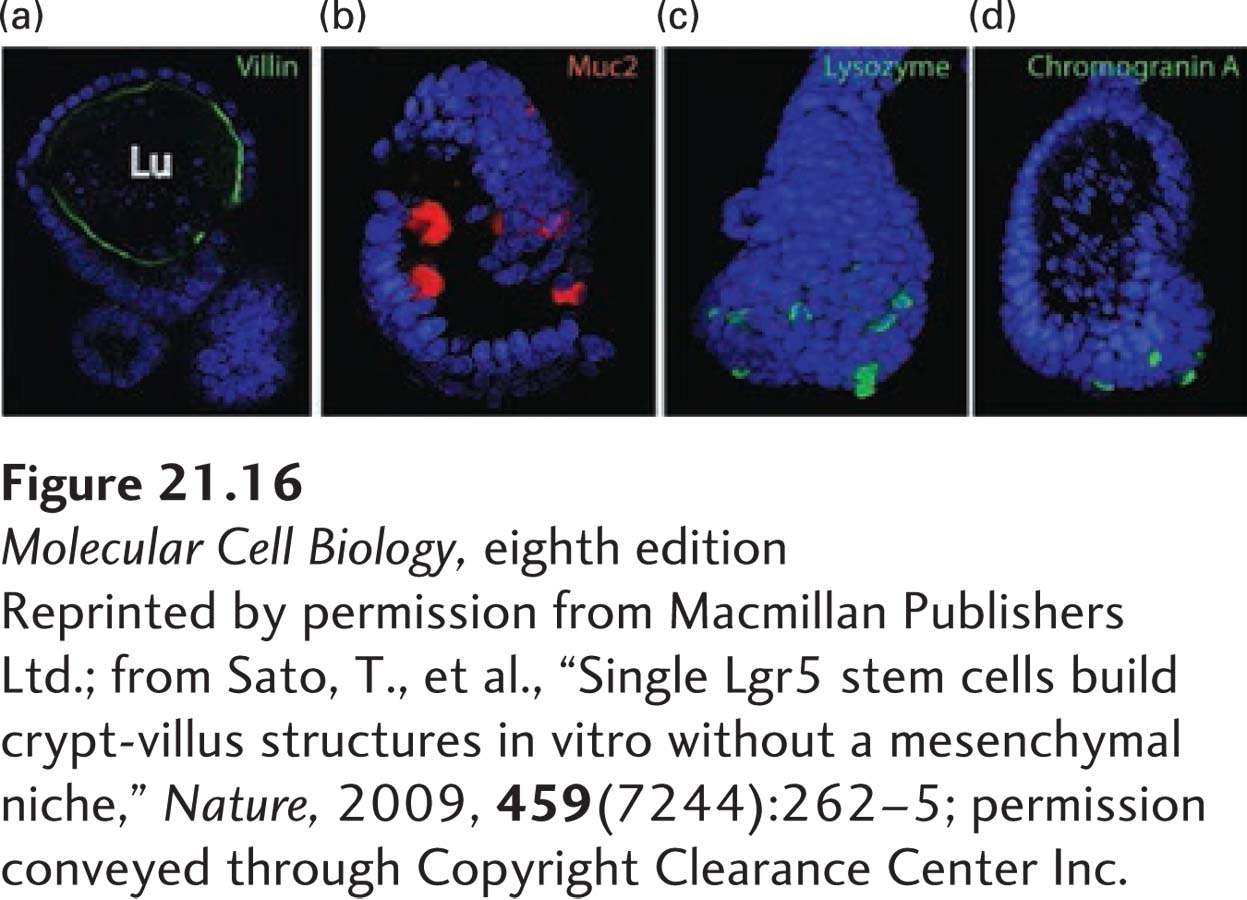
In subsequent studies, single Lgr5-expressing stem cells were isolated from intestinal crypts and cultured on an extracellular matrix (see Figure 20-23) containing type IV collagen and laminin, similar to the matrix that normally underlies and supports the intestinal epithelia. These cells generated villus-like structures that contained all four differentiated cell types found in the mature intestinal epithelium (Figure 21-16). Taken together, these experiments established that expression of the Lgr5 gene defines the intestinal stem cells and showed that these cells are localized at the bases of the intestinal crypts interspersed between the terminally differentiated Paneth cells (see Figure 21-13). Lgr5-expressing cells are also found in the stomach, colon, and pancreas—which, like the small intestine, are formed from the embryonic endoderm—and are thought to be the stem cells for these tissues. Indeed, culturing Lgr5-expressing cells from these tissues in the presence of Wnt, R-spondin, and other hormones generates mini-organoids that contain differentiated cells characteristic of these tissues.
[Reprinted by permission from Macmillan Publishers Ltd.; from Sato, T., et al., “Single Lgr5 stem cells build crypt-villus structures in vitro without a mesenchymal niche,” Nature, 2009, 459(7244):262–5; permission conveyed through Copyright Clearance Center Inc.]
EXPERIMENTAL FIGURE 21-16Single Lgr5-expressing intestinal stem cells build crypt-villus structures in culture without niche cells. Single Lgr5-expressing cells isolated from intestinal crypts were placed in culture on a type IV extracellular matrix (see Figure 20-23), the type of matrix that normally underlies and supports sheets of epithelial cells. After 2 weeks, these cultures had formed epithelial sheets that resembled villi in structure. Staining of these organoids for specific marker proteins showed that they contained all four differentiated epithelial cell types: (a) villin (green) is a marker protein for the absorptive enterocytes that are localized near the apical (luminal, Lu) surface of these organoids; (b) Muc2 (red) for goblet cells; (c) lysozyme (green) for Paneth cells; and (d) chromogranin A (green) for enteroendocrine cells. The organoids were also stained with DAPI (blue) to reveal nuclei.
[Reprinted by permission from Macmillan Publishers Ltd.; from Sato, T., et al., “Single Lgr5 stem cells build crypt-villus structures in vitro without a mesenchymal niche,” Nature, 2009, 459(7244):262–5; permission conveyed through Copyright Clearance Center Inc.]
Page 993
Page 994
Paneth cells are longer-lived than the cells of the villi; they produce several antibacterial proteins, including the enzyme lysozyme, which degrades bacterial cell walls and thus protects the intestine from infections. Surprising recent evidence suggests that Paneth cells also constitute a major part of the niche for the intestinal stem cells. Cultured Paneth cells produce Wnt as well as other hormones, such as EGF and a Delta protein (see Chapter 16), that are essential for intestinal stem-cell maintenance. Co-culturing of intestinal stem cells with Paneth cells markedly improved the formation of intestinal villus-like structures, and genetic manipulations in mice that caused a reduction of Paneth cell numbers concomitantly caused a reduction in intestinal stem cells. Thus Paneth cells—which are progeny cells of the intestinal stem cells—constitute much, if not all, of the niche for intestinal stem-cell maintenance.
The Lgr5-expressing cells may not be the only type of intestinal stem cells. Evidence indicates that so-called +4 cells located in the crypts (see Figure 21-13a) may be “reserve stem cells” that can generate Lgr5-expressing stem cells following intestinal injury, such as by irradiation. In turn, these +4 cells can be generated from Lgr5-expressing stem cells (see Figure 21-13b). Recall that transient amplifying cells have limited self-renewal potential (see Figure 21-10). During periods of intestinal injury, when many Lgr5-expressing stem cells are lost, some of the transient amplifying cells, under the influence of Wnt signals, can “dedifferentiate” and revert to Lgr5-expressing stem cells and relocalize to the Paneth-cell niche! Thus the conversion of differentiated cells into stem cells, as seen experimentally during formation of iPS cells, may occur normally in the body during periods of stress or injury. Thus, rather than relying on a single stem cell-to-differentiated cell lineage (see Figure 21-13b), the intestine seems capable of drawing on several pools of reserve stem-cell populations. Future work will undoubtedly clarify the role of these cells in generation of intestinal epithelial cells.