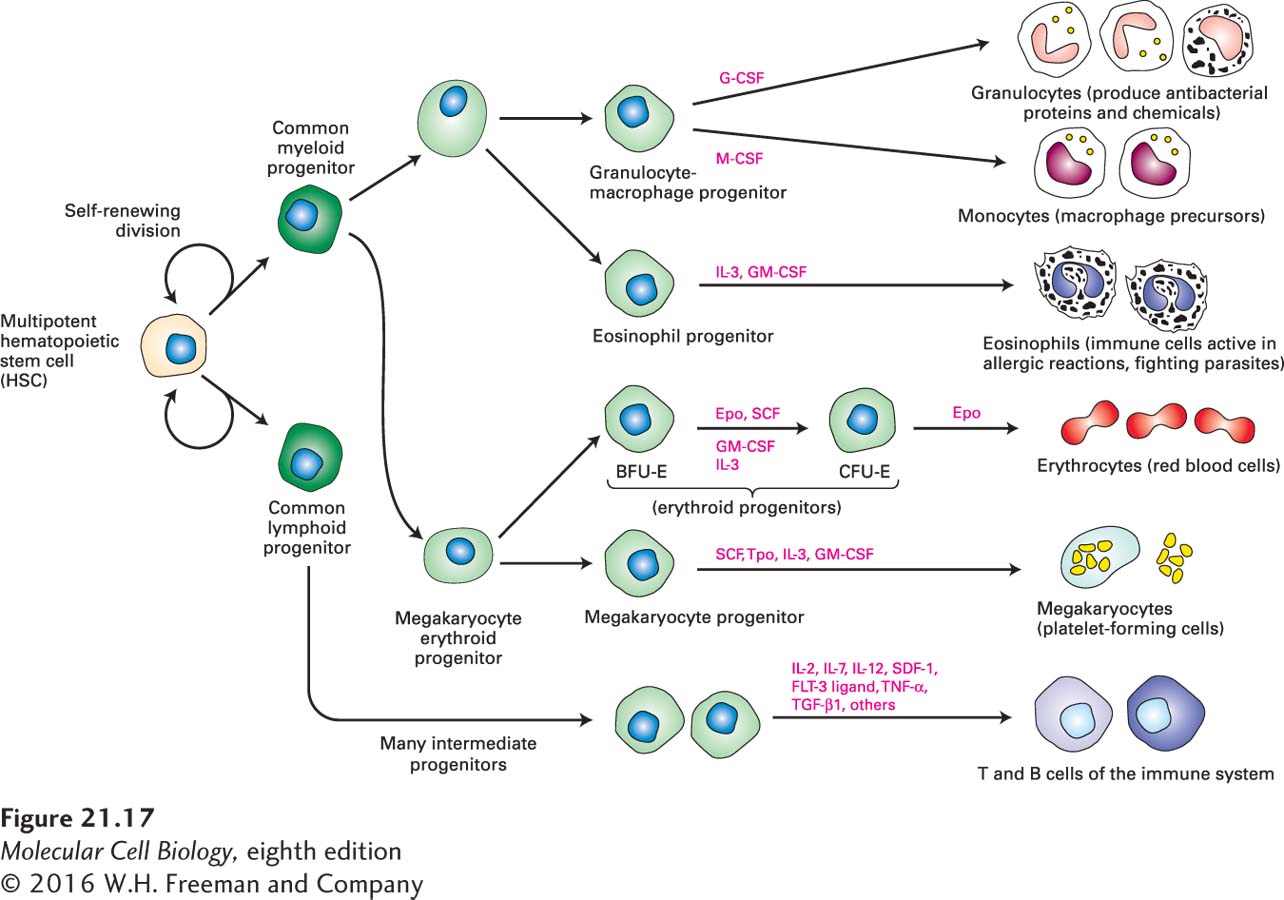
Another continuously replenished tissue is the blood, whose stem cells are located in the embryonic liver and in bone marrow in adult animals. The various types of blood cells all derive from a single type of multipotent, self-renewing hematopoietic stem cell (HSC). An HSC gives rise to two other multipotent cell types, common myeloid and common lymphoid progenitor cells, which are more restricted in their fates but are capable of limited self-renewal (Figure 21-17). Numerous extracellular growth factors called cytokines regulate HSC self-renewal divisions as well as proliferation and differentiation of the precursor cells for various blood-cell lineages. Each branch of the blood-cell lineage tree has different cytokine regulators, allowing exquisite control of the production of specific cell types. If all blood-cell types are needed—for example, after a bleeding injury—multiple cytokines can be produced. If only one cell type is needed, specific signals control its production. For example, when a person is traveling at high altitude, erythropoietin is made by the kidney and stimulates the proliferation and differentiation of CFU-E (erythroid progenitor) cells, but not other types of blood-cell precursors. Erythropoietin activates several different intracellular signal transduction pathways, leading to changes in gene expression that promote formation of erythrocytes (see Figures 16-7 and 16-8). Similarly, G-CSF, a different cytokine, stimulates proliferation of bipotential granulocyte-macrophage progenitors and their differentiation into granulocytes, while M-CSF stimulates production of macrophages from the same progenitor cell type.
FIGURE 21-17Formation of blood cells from hematopoietic stem cells in the bone marrow. Multipotent hematopoietic stem cells may divide symmetrically to increase the numbers of stem cells. In adults, they generally divide asymmetrically to form one daughter cell that is multipotent, like the parent stem cell, and another daughter cell with a more restricted fate. Ultimately, this daughter cell generates either common lymphoid progenitors or common myeloid progenitors; although these multipotent cells are capable of limited self-renewal, they are committed to one of the two major hematopoietic lineages. Depending on the types and amounts of cytokines present, the common lymphoid and common myeloid progenitors undergo rapid rounds of cell division and generate different types of progenitor cells (light green). These progenitors are either multipotent or unipotent in that they can give rise to several types or only a single type of differentiated blood cells, respectively; they respond to one or a few specific cytokines. Some of the cytokines that support this process are indicated (pink labels). CSF = colony-stimulating factor; IL = interleukin; SCF = stem-cell factor; Epo = erythropoietin; Tpo = thrombopoietin. See M. Socolovsky et al., 1998, Proc. Natl. Acad. Sci. USA95:6573, and N. Noverstern et al., 2011, Cell144:296.
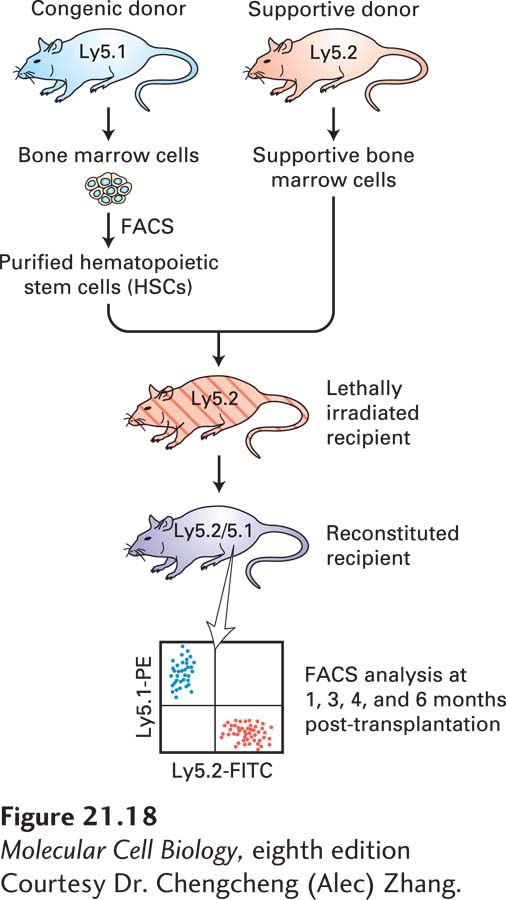
Hematopoietic stem cells were originally detected and quantified by bone marrow transplantation experiments in mice whose hematopoietic stem and progenitor cells had been wiped out by irradiation (Figure 21-18). By transplanting specific types of hematopoietic precursors into these mice and observing which blood cells were restored, researchers could infer which precursors or terminally differentiated cells (e.g., erythrocytes, monocytes) arise from a particular type of precursor. The first step was separation of the different types of precursors. This sorting was possible because HSCs and each type of precursor produce unique combinations of cell-surface proteins that can serve as cell type–specific markers. If bone marrow extracts are treated with fluorochrome-labeled antibodies for these markers, cells with different markers can be separated in a fluorescence-activated cell sorter (FACS; see Figures 4-2 and 4-3). Remarkably, such transplantation experiments revealed that a single HSC is sufficient to restore the entire blood system when transferred into a lethally irradiated mouse in which all of the HSCs have been killed. After transplantation, the HSC takes up residence in a niche in the bone marrow and divides to make more HSCs as well as progenitors of the different blood-cell lineages.
[Courtesy Dr. Chengcheng (Alec) Zhang.]
EXPERIMENTAL FIGURE 21-18Functional analysis of hematopoietic stem cells by bone marrow transplantation. The two strains of mice used in this analysis are genetically identical except for the gene encoding a protein, termed Ly5, found on the surfaces of all nucleated blood cells, including all T and B lymphocytes, granulocytes, and monocytes. The proteins encoded by the two alleles of the gene, Ly5.1 and Ly5.2, can be detected by specific monoclonal antibodies. A recipient Ly5.2 mouse is lethally irradiated to kill all HSCs, then injected with stem cells purified from a Ly5.1 strain. Because the stem cells take weeks or months to produce differentiated blood cells, the recipient mouse will die unless it receives bone marrow progenitor cells from a genetically identical mouse (termed “supportive” cells) that will produce mature blood cells for the first few weeks after the transplant. At intervals after the transplant, blood or bone marrow is recovered and reacted with a blue-fluorescing monoclonal antibody to Ly5.1 and a red-fluorescing monoclonal antibody to Ly5.2. Mature blood cells that are descended from the donor stem cell are detected by FACS analysis, seen here as cells that fluoresce blue and not red. These cells can be sorted and stained with fluorescent antibodies specific for marker proteins found on different types of mature blood cells to show that a stem cell is indeed pluripotent, in that it can generate all types of lymphoid and myeloid cells.
[Courtesy Dr. Chengcheng (Alec) Zhang.]
Page 995
The first successful human bone marrow transplant was done in 1959, when a patient with end-stage (fatal) leukemia was irradiated to destroy her cancer cells as well as her own normal HSCs. She was transfused with bone marrow cells from her identical twin sister, thus avoiding an immune response, and was in remission for 3 months. This pioneering effort, which was awarded the Nobel Prize in Medicine in 1990, led to the present-day treatments that can often lead to a complete cure of many cancers. The stem cells in the transplanted marrow can generate all types of functional blood cells, so transplants are useful in patients with certain hereditary blood diseases, including many genetic anemias (insufficient red-cell levels) or genetic defects of blood cells, such as sickle-cell disease (a hemoglobin disorder), as well as in cancer patients who have received irradiation or chemotherapy, both of which destroy the bone marrow cells as well as cancer cells.
During embryonic life, HSCs often divide symmetrically, producing two daughter stem cells (see Figure 21-11); this process allows the number of stem cells to increase over time and produce the large number of progenitor cells required to make all of the necessary blood cells before birth. In adult animals, HSCs are largely quiescent, “resting” in the G0 state in the bone marrow stem-cell niche. When more blood cells are needed, cytokines are generated that signal HSCs to divide, producing stem cells like the parent cells and rapidly proliferating transient amplifying cells that generate the progenitors illustrated in Figure 21-17. Whether individual HSCs undergo symmetric or asymmetric division is not known.