Meristems Are Niches for Stem Cells in Plants
![]() In plants, as in their multicellular animal counterparts, the production of all tissues and organs relies on small populations of stem cells. Like animal stem cells, these stem cells are defined by their ability to undergo self-
In plants, as in their multicellular animal counterparts, the production of all tissues and organs relies on small populations of stem cells. Like animal stem cells, these stem cells are defined by their ability to undergo self-
Page 997
The niches in which plant stem cells are located, called meristems, can persist for thousands of years in long-
Page 998
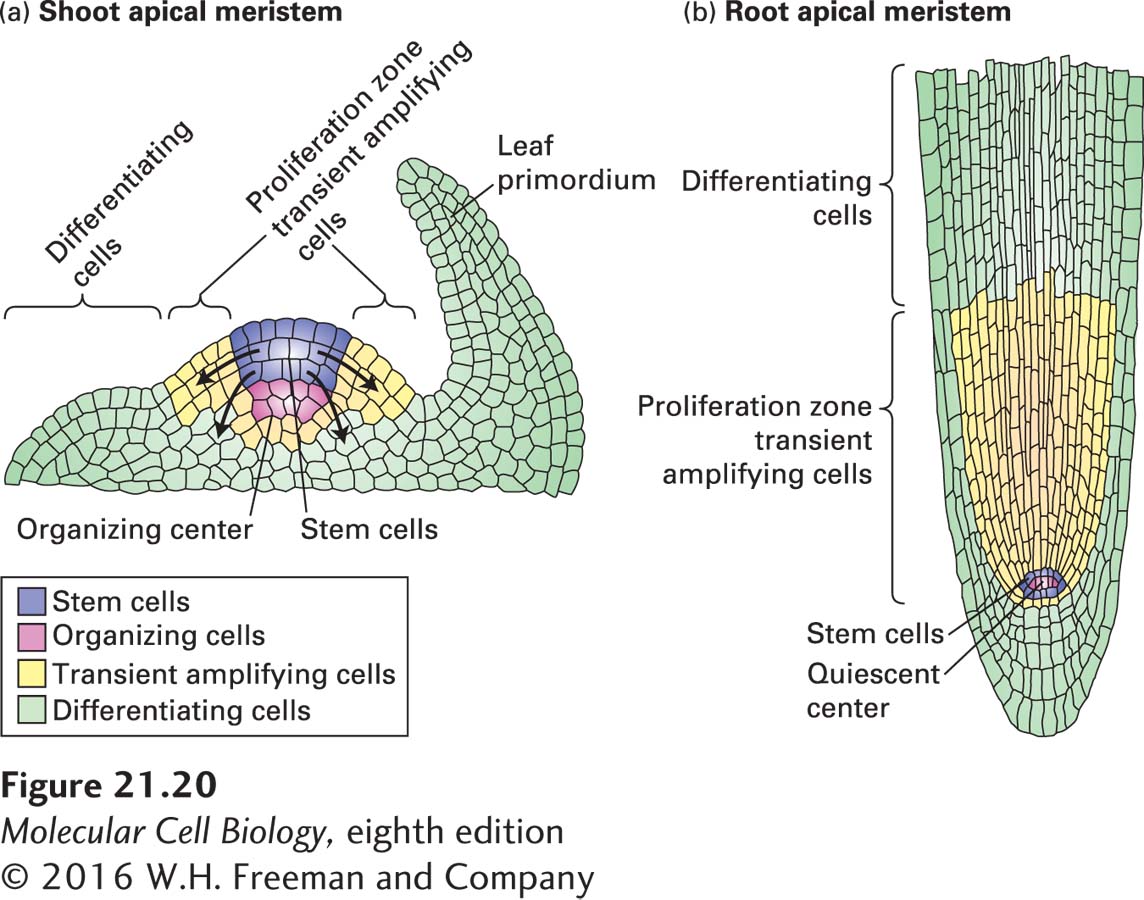
Unlike somatic stem cells in metazoan animals, somatic plant stem cells give rise to entire organs, not just specific tissues or lineages. Slowly dividing pluripotent stem cells are located at the apex of the shoot apical meristem, with more rapidly dividing multipotent transient amplifying daughter cells on the periphery. Descendants of the shoot stem cells are displaced to the periphery of the meristem and are recruited to form primordia of new organs, including leaves and stems. Division ceases as these cells acquire the characteristics of specific cell types, and most organ growth occurs by cell expansion and elongation (Figure 21-20a). New shoot stem-