The Par Proteins and Other Polarity Complexes Are Involved in Epithelial-Cell Polarity
In vertebrates, polarized epithelial cells use cues from adjacent cells and the extracellular matrix to orient their axis of polarization. The process of polarization in epithelial cells of vertebrates is quite similar to that in the fruit fly Drosophila melanogaster. Much of our knowledge has come from the fly system because of the ease with which mutants can be isolated and analyzed.
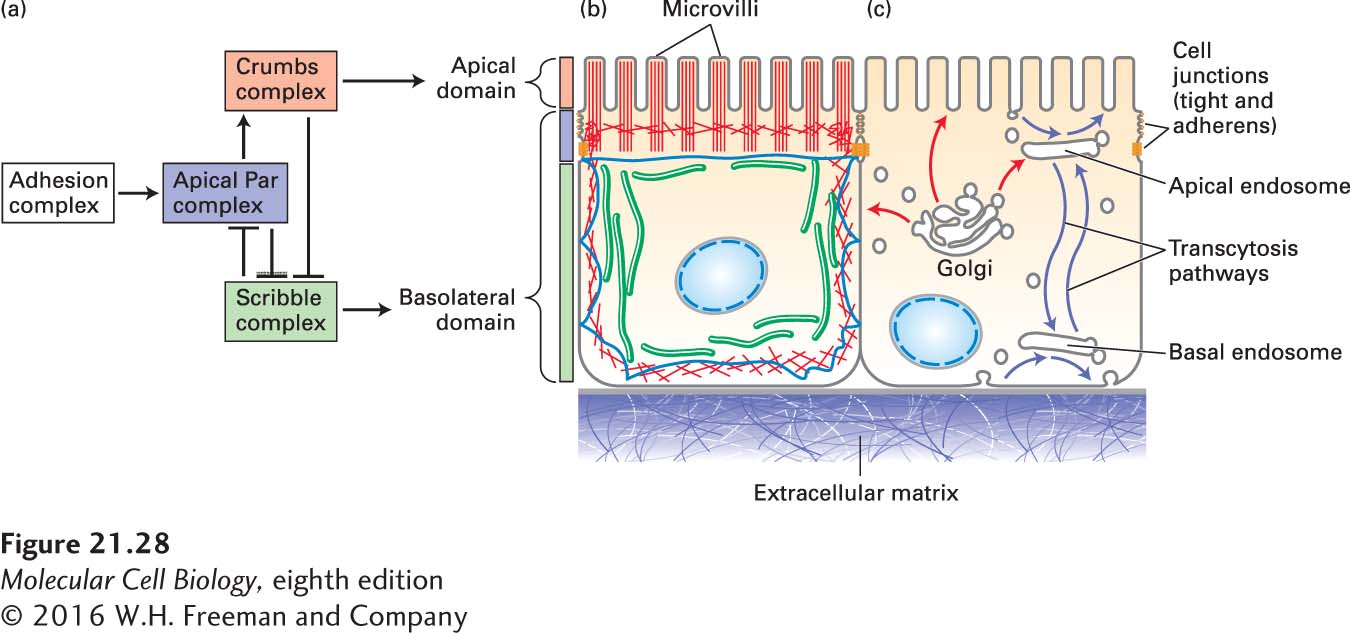
Genetic screens in the fly have uncovered multiple genes necessary for the generation of epithelial-cell polarity. Analyses of the proteins encoded by these genes and of the phenotypes of mutants have identified three major groups of proteins: the complex made up of Cdc42, Par3, Par6, and aPKC (in this system known as the apical Par complex, or simply as the Par complex), the Crumbs complex, and the Scribble complex. By extensive analyses of the effects of these complexes on one another when individual components are missing, a general understanding of their contributions to epithelial-cell polarization has been achieved, although a detailed molecular understanding is still emerging (Figure 21-28a).
FIGURE 21-28Establishment of polarity in epithelial cells. (a) Polarity determination in epithelial cells, like that in the nematode embryo, is driven by an apical Par complex. The formation of a cell-cell adhesion complex induces the recruitment of the Par complex. Then, intricate and antagonistic interactions of the Par complex with both the basolateral Scribble complex and the apical Crumbs complex lead to the establishment and maintenance of epithelial-cell polarity. The localization of the different complexes to membrane domains is indicated by colored bars: the Scribble complex associates with the lateral membrane, the Par complex associates with the region at the cell junctions, and the Crumbs complex is immediately apical to the Par complex. Functional epithelial polarity is maintained by both (b) a polarized cytoskeleton and (c) membrane trafficking pathways. In the biosynthetic pathway, proteins and lipids destined for the apical and basolateral domains are sorted in the Golgi complex and transported to their respective surfaces (red arrows). Endocytic pathways (blue arrows) regulate the abundances of proteins and lipids on each surface and sort them between surfaces by transcytosis.
The first known step in epithelial-cell polarization is interaction between adjacent cells, which in vertebrate cells occurs through nectin, a cell-adhesion molecule in the Ig superfamily, and a junctional protein called JAM-A. These interactions signal the cells to recruit the Par complex and to assemble adherens and tight junctions (see Figure 20-1). The Crumbs complex is recruited more apically than the Par complex, and the Scribble complex defines the basolateral surface. In the absence of the Par complex, cells cannot polarize, and as in the nematode embryo, the Par complex is the master regulator of cell polarity. In the absence of the Scribble complex, the apical domain is greatly expanded, whereas in the absence of Crumbs, the apical domain is greatly reduced. These observations have led to the realization that there are mutually antagonistic relationships between these complexes, in which, for example, the apical Par complex kinase aPKC antagonizes the basolateral Scribble complex by phosphorylation (see Figure 21-28a). Thus, as is the case in the nematode embryo, asymmetry is mediated by complexes working antagonistically against each other.
In a manner that is only partially understood, this arrangement of polarity proteins reorganizes the cytoskeleton so that distinct arrangements of microfilaments become associated with the apical and basolateral membranes. The distribution of microtubules in epithelial cells is rather unusual, as they do not all associate with a centrosome; instead, lateral microtubules orient their (−) ends toward the apical domain and other microtubules run perpendicular to the lateral microtubules below the microvilli and also along the base of the cell (Figure 21-28b); how these arrangements are established is not known. Membrane traffic is also polarized (Figure 21-28c). Newly made membrane proteins destined for the apical and basolateral membranes are sorted and packaged into distinct transport vesicles at the trans-Golgi network and then transported to the appropriate surface. In addition, endocytic pathways from both the apical and basolateral surfaces regulate the abundance of membrane proteins and transport missorted proteins using a complex set of sorting endosomes in a process known as transcytosis.
Page 1008
In genetic screens for additional components important for epithelial-cell polarity in the fly, components of endocytic trafficking were found. For example, one such mutant affects the trafficking of the apical transmembrane protein Crumbs, so that when endocytosis is compromised, the level of Crumbs on the surface goes up and the apical domain expands. Thus epithelial-cell polarity involves responses to spatial cues and reorganization of the cytoskeleton that provides a framework for both secretory and endocytic membrane traffic pathways for establishment and maintenance of the polarized state.