Evolutionarily Conserved Proteins Participate in the Apoptotic Pathway
The confluence of genetic studies in C. elegans and studies on human cancer cells suggested that an evolutionarily conserved pathway mediates apoptosis. In C. elegans, cell lineages are under tight genetic control and are identical in all individuals of the species. About 10 rounds of cell division or fewer create the adult worm, which is about 1 mm long and 70 µm in diameter. The adult worm may be a hermaphrodite (a worm with both male and female organs) or a male. The hermaphrodite form has 959 somatic-cell nuclei, whereas the male has 1031 (see Figure 21-25d). Scientists have traced the lineage of each somatic cell in C. elegans from the fertilized egg to the mature worm by following the development of live worms using DIC microscopy (see Figure 21-25c).
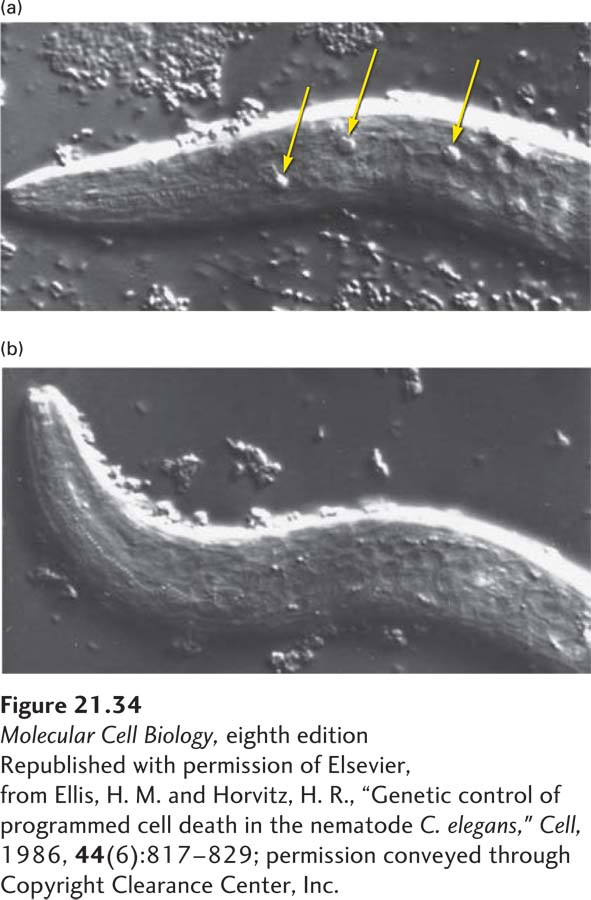
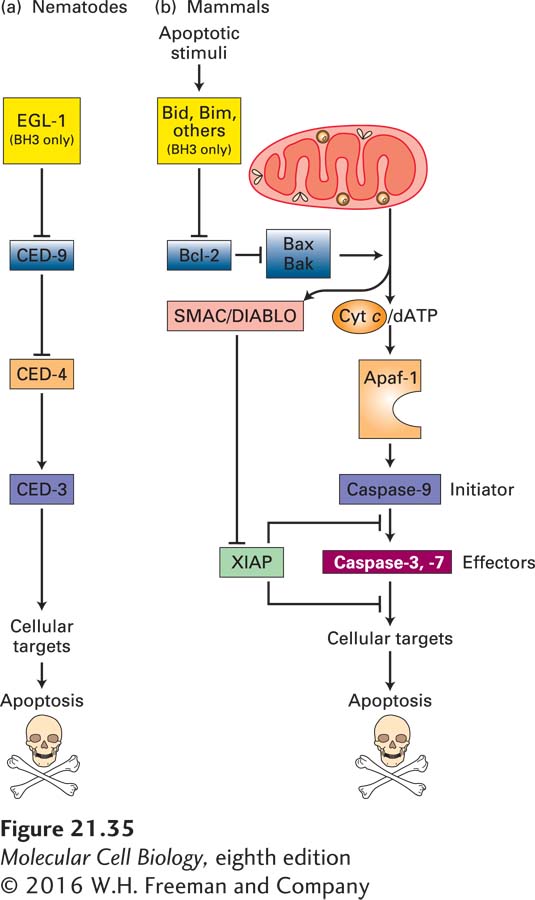
Of the 947 nongonadal cells generated during development of the adult hermaphrodite form, 131 undergo programmed cell death. Specific mutations have revealed four genes whose encoded proteins play an essential role in controlling programmed cell death during C. elegans development: ced-3, ced-4, ced-9, and egl-1. In ced-3 or ced-4 mutants, for example, the 131 “doomed” cells survive (Figure 21-34). These mutants formed the first pieces of evidence that apoptosis was under a genetic program and led to a Nobel Prize for Robert Horvitz. The mammalian proteins that correspond most closely to the worm CED-3, CED-4, CED-9, and EGL-1 proteins are indicated in Figure 21-35. In discussing the worm proteins, we will occasionally include the mammalian names in parentheses to make it easier to keep the relationships clear.
[Republished with permission of Elsevier, from Ellis, H. M. and Horvitz, H. R., “Genetic control of programmed cell death in the nematode C. elegans,” Cell, 1986, 44(6):817–829; permission conveyed through Copyright Clearance Center, Inc.]
EXPERIMENTAL FIGURE 21-34Mutations in the ced-3 gene block programmed cell death in C. elegans. (a) Newly hatched mutant larva carrying a mutation in the ced-1 gene. Because mutations in this gene prevent engulfment of dead cells, highly refractile (and thus easily visualized) dead cells accumulate (arrows). (b) Newly hatched larva with mutations in both the ced-1 and ced-3 genes. The absence of refractile dead cells in these double mutants indicates that no cell deaths occurred. Thus the CED-3 protein is required for programmed cell death.
[Republished with permission of Elsevier, from Ellis, H. M. and Horvitz, H. R., “Genetic control of programmed cell death in the nematode C. elegans,” Cell, 1986, 44(6):817–829; permission conveyed through Copyright Clearance Center, Inc.]
FIGURE 21-35Evolutionary conservation of apoptosis pathways. Similar proteins, shown in identical colors, play corresponding roles in nematodes and in mammals. (a) In nematodes, the BH3-only protein called EGL-1 binds to CED-9 on the outer mitochondrial membrane; this interaction releases CED-4 from the CED-4/CED-9 complex. Free CED-4 then binds to, and activates by autoproteolytic cleavage, the caspase CED-3, which destroys cell proteins to drive apoptosis. These relationships are shown as a genetic pathway, with EGL-1 inhibiting CED-9, which in turn inhibits CED-4. Active CED-4 activates CED-3. (b) In mammals, homologs of the nematode proteins, as well as many other proteins not found in the nematode, regulate apoptosis. The Bcl-2 protein is similar to CED-9 in promoting cell survival. It does so in part by preventing activation of Apaf-1, which is similar to CED-4, and in part by other mechanisms depicted in Figure 21-40. Several types of BH3-only proteins, detailed in Figures 21-39 and 21-40, inhibit Bcl-2 and thus allow apoptosis to proceed. Many apoptotic stimuli lead to damage of the outer mitochondrial membrane, causing release into the cytosol of several proteins that stimulate apoptosis. In particular, cytochrome c released from mitochondria activates Apaf-1, which in turn activates caspase-9. This initiator caspase then activates effector caspases-3 and -7, eventually leading to apoptosis. See text for discussion of other mammalian proteins (SMAC/DIABLO and XIAP) that have no nematode homologs. See S. J. Riedl and Y. Shi, 2004, Nat. Rev. Mol. Cell Biol.5:897.
The first mammalian apoptotic gene to be cloned, bcl-2, was isolated from human follicular lymphomas, tumors of the antibody-producing B cells of the immune system. A mutant form of this gene was formed in a patient’s lymphoma cells; a chromosomal rearrangement joined the protein-coding region of the bcl-2 gene to an immunoglobulin-gene enhancer. The combination results in overproduction of the Bcl-2 protein, which keeps these cancer cells alive when they would otherwise become programmed to die. The human Bcl-2 protein and worm CED-9 protein are homologous; even though the two proteins are only 23 percent identical in sequence, expression of a bcl-2 transgene can block the extensive cell death observed in ced-9 mutant worms. Thus both proteins act as regulators that suppress the apoptotic pathway (see Figure 21-35). In addition, both proteins contain a single transmembrane domain and are localized mainly to the outer mitochondrial membrane, where they serve as sensors that control the apoptotic pathway in response to external stimuli. As we discuss next, other regulators promote apoptosis.
In the worm apoptotic pathway, CED-3 (caspase-9 in mammals) is a protease required to destroy cell components during apoptosis. CED-4 (Apaf-1) is a protease-activating factor that causes autoproteolytic cleavage of (and by) the inactive precursor of CED-3 (a zymogen), creating an active CED-3 protease that initiates cell death (see Figures 21-35 and 21-36). Cell death does not occur in ced-3 and ced-4 single mutants or in ced-9/ced-3 double mutants. In contrast, in ced-9 mutants, all cells die by apoptosis during embryonic life, so the adult form never develops. These genetic studies indicate that CED-3 and CED-4 are killer proteins required for cell death, and that CED-9 (Bcl-2) suppresses apoptosis. The observation that all cells die in ced-9 mutants shows that the apoptotic pathway is present in and can be activated in all body cells. Moreover, the absence of cell death in ced-9/ced-3 double mutants suggests that CED-9 acts upstream of CED-3 to suppress the apoptotic pathway.
Page 1014
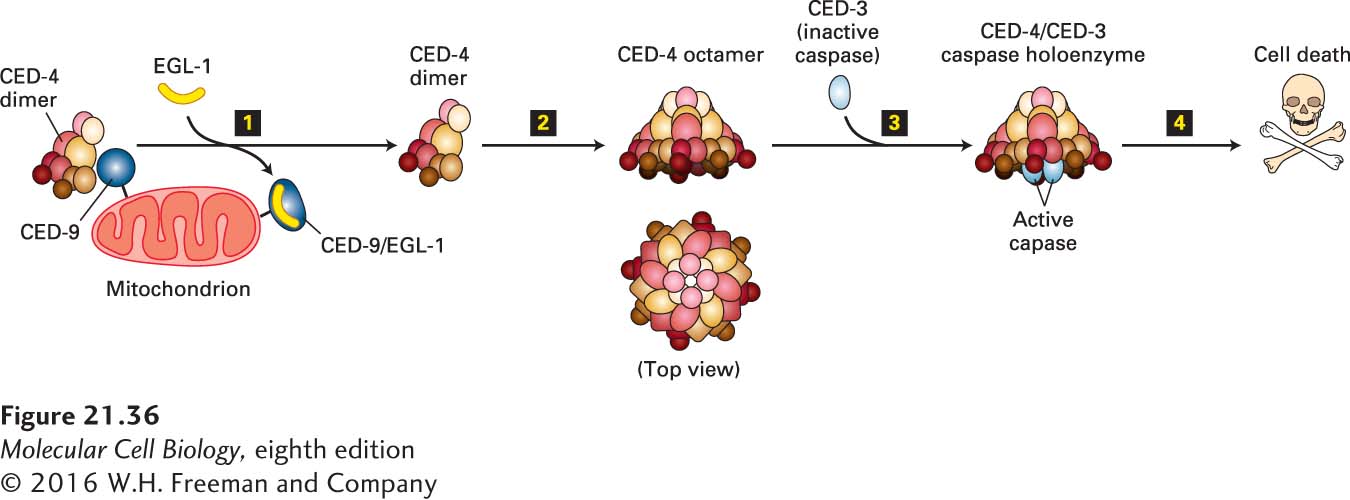
The mechanism by which CED-9 (Bcl-2) controls CED-3 (caspase-9) in the nematode is known and is somewhat different from the mechanism, discussed later (see Figure 21-41), in mammalian cells. The nematode CED-9 protein forms a complex with a dimer of the CED-4 (Apaf-1) protein, thereby preventing the activation of CED-3 by CED-4 (Figure 21-36). As a result, the cell survives. This mechanism fits with the genetics, which shows that the absence of CED-9 has no effect if CED-3 is also missing (ced-3/ced-9 double mutants have no cell death). The three-dimensional structure of the trimeric CED-4/CED-9 complex reveals a huge contact surface between each of the two CED-4 molecules and the single CED-9 molecule; the large contact surface makes the association highly specific, but in such a way that the dissociation of the complex can be regulated.
FIGURE 21-36Activation of CED-3 protease in C. elegans. EGL-1 protein, which is produced in response to developmental signals that trigger cell death, displaces an asymmetric CED-4 dimer from its association with CED-9 (step 1). The free CED-4 dimer combines with three others to form an octamer (step 2), which binds two molecules of the CED-3 zymogen (an enzymatically inactive precursor of a caspase protease) and triggers the conversion of the CED-3 zymogen into active CED-3 protease (step 3). This effector caspase then begins to destroy cell components, leading to cell death (step 4). See N. Yan et al., 2005, Nature437:831, and S. Qi et al., 2010, Cell141:446.
Transcription of egl-1, the fourth genetically defined apoptosis regulator gene, is stimulated in C. elegans cells that are programmed to die. Newly produced EGL-1 protein binds to CED-9, alters its conformation, and catalyzes the release of CED-4 from CED-9 (see Figure 21-36). Both EGL-1 and CED-9 contain a 12-amino-acid BH3 domain. Because EGL-1 lacks most of the other domains of CED-9, it is called a BH3-only protein. The mammalian BH3-only proteins closest in sequence and function to EGL-1 are the pro-apoptotic proteins Bid and Bim, discussed later.
Insight into how EGL-1 disrupts the CED-4/CED-9 complex comes from the molecular structure of EGL-1 (mammalian Bid/Bim) complexed with CED-9 (Bcl-2). In this complex, the BH3 domain forms the key part of the contact surface between the two proteins. CED-9 has a different conformation when bound by EGL-1 than when bound by CED-4. This finding suggests that EGL-1 binding distorts CED-9, making its interaction with CED-4 less stable. Once EGL-1 causes dissociation of the CED-4/CED-9 complex, the released CED-4 dimer joins with three other CED-4 dimers to make an octamer, which then activates CED-3 by a mechanism we will discuss shortly. Cell death soon follows (see Figure 21-36).
Evidence that the steps described here are sufficient to induce apoptosis comes from experiments in which these steps were reconstituted in vitro with purified proteins. CED-3, CED-4, a segment of the CED-9 protein that lacked its mitochondrial membrane anchor, and EGL-1 were purified, as was a CED-4/CED-9 complex. Purified CED-4 (Apaf-1) was able to accelerate the autoproteolytic cleavage and activation of purified CED-3 (caspase-9), but addition of CED-9 (Bcl-2) to the reaction mixture inhibited the autocleavage. When the CED-4/CED-9 complex was mixed with CED-3, autocleavage did not occur, but addition of EGL-1 to the reaction mixture restored CED-3 autocleavage by releasing CED-4 from its complex with CED-9.
Page 1015
To see the importance of regulated EGL-1 expression in apoptosis, consider a class of neurons in C. elegans found in hermaphrodites, but not in males. These hermaphrodite-specific neurons are generated embryonically in both hermaphrodites and males, but undergo programmed cell death in males. In hermaphrodites, expression of the egl-1 gene in these neurons is repressed by the transcription factor TRA-1A, and deletion of TRA-1A in hermaphrodites causes these neurons to undergo apoptosis. This finding reinforces a point made earlier: all metazoan cells can potentially undergo apoptosis, so this process needs to be carefully regulated!