Cleavage of the Mammalian Embryo Leads to the First Differentiation Events
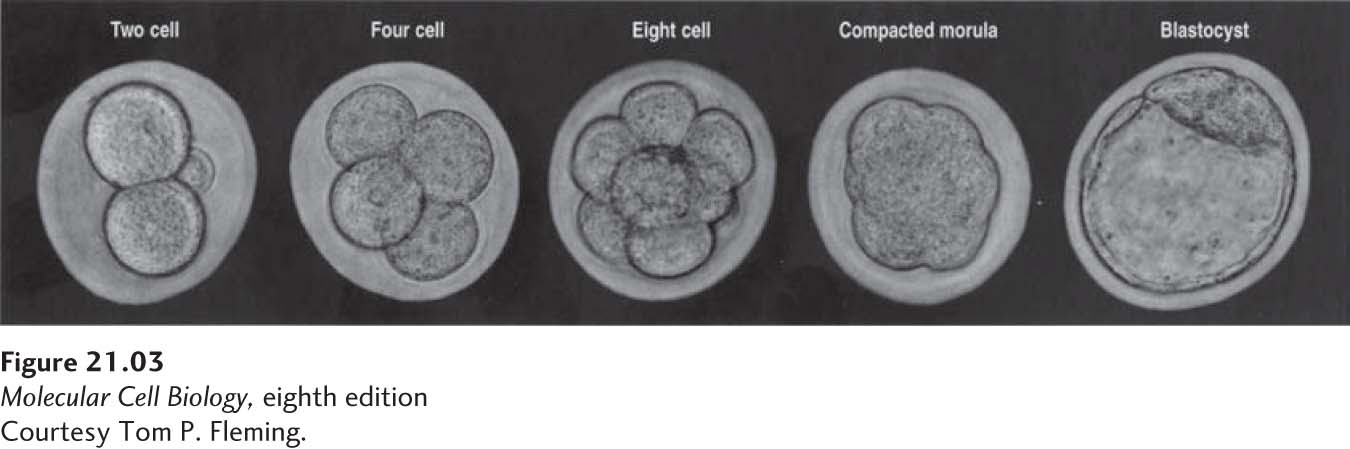
The fertilized egg, or zygote, does not remain a single cell for long. Fertilization is quickly followed by cleavage, a series of cell divisions that take about one day each (Figure 21-3); these divisions happen before the embryo is implanted in the uterine wall. Initially, the cells are fairly spherical and loosely attached to one another. As demonstrated experimentally in sheep, each cell at the 8-cell stage is totipotent and has the potential to give rise to a complete animal when implanted into the uterus of a pseudopregnant animal (one treated with hormones to make her uterus responsive to embryos).
[Courtesy Tom P. Fleming.]
FIGURE 21-3Cleavage divisions in the mouse embryo. There is little cell growth during these early divisions, so that the cells become progressively smaller. See text for discussion.
[Courtesy Tom P. Fleming.]
Three days after fertilization, the 8-cell embryo divides again to form the 16-cell morula (from the Greek for “raspberry”), after which the cell affinities for one another increase substantially and the embryo undergoes compaction, a process that depends in part on the cell-surface homotypic cell-adhesion protein E-cadherin (see Figure 20-14). The compaction process driven by increased cell-cell adhesion initially results in a more solid mass of cells, the compacted morula. In the next step, some of the cell-cell adhesions diminish locally, and fluid begins to flow into an internal cavity called the blastocoel. Additional divisions produce a blastocyst (see Figure 21-3).
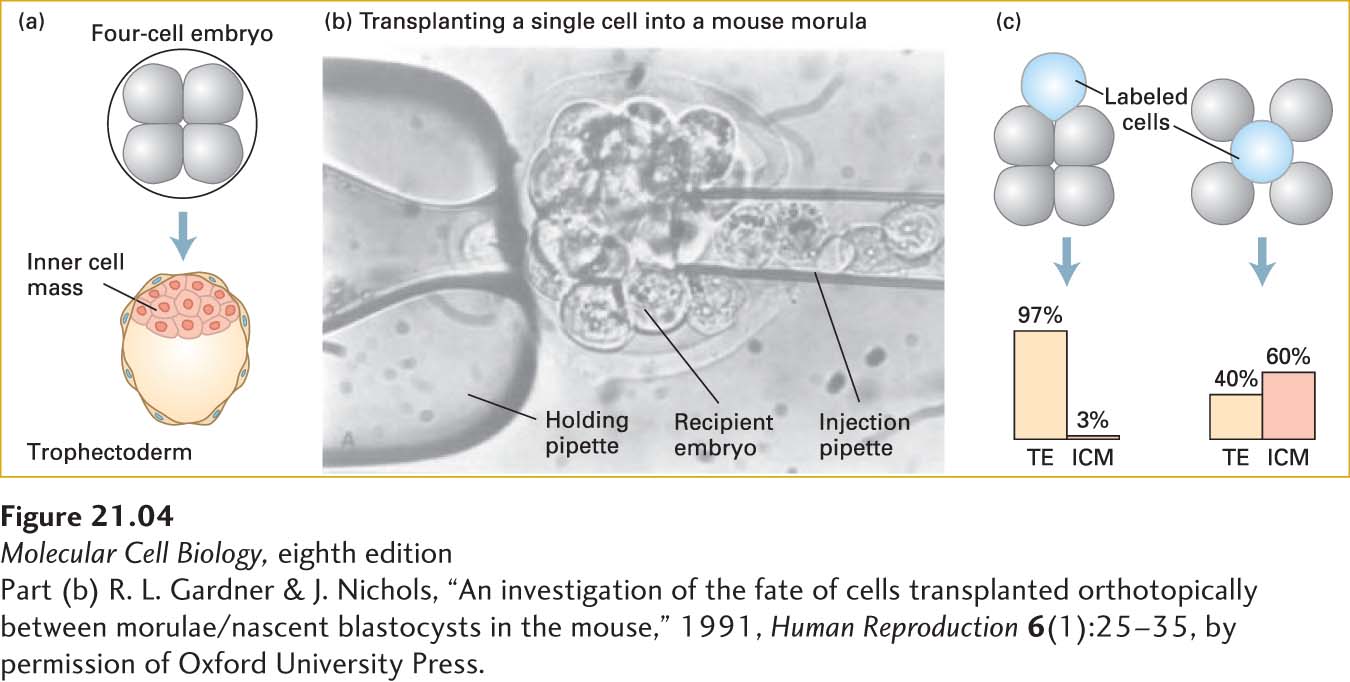
The blastocyst is composed of approximately 64 cells that have separated into two cell types: trophectoderm (TE), which will form extraembryonic tissues such as the placenta, and the inner cell mass (ICM) (just 10–15 cells in a mouse), which gives rise to the embryo proper (Figure 21-4a). In the blastocyst, the ICM is found on one side of the blastocoel, while the TE cells form a hollow ball around the ICM and blastocoel. At this point, the TE cells are in an epithelial sheet, while the ICM cells are a loose mass that can be described as mesenchyme. Mesenchyme, a term most commonly applied to mesoderm-derived cells, refers to loosely organized and loosely attached cells.
[Part (b) R. L. Gardner & J. Nichols, “An investigation of the fate of cells transplanted orthotopically between morulae/nascent blastocysts in the mouse,” 1991, Human Reproduction6(1):25–35, by permission of Oxford University Press.]
EXPERIMENTAL FIGURE 21-4Cell location determines cell fate in the early embryo. (a) A four-cell embryo normally develops into a blastocyst consisting of trophectoderm (TE) cells on the outside and inner cell mass (ICM) cells inside. (b) In order to discover whether position affects the fates of cells, transplantation experiments were done with mouse embryos. First, recipient morula-stage embryos had cells removed to make room for implanted cells. Then donor morula-stage (sixteen-cell) embryos were soaked in a dye that does not transfer between cells. Finally, labeled cells from the donor embryos were injected into inner or outer regions of the recipient embryos, as shown in the micrograph. The recipient embryo was held in place by a slight vacuum applied to the holding pipette. (c) The subsequent fates of the descendants of the transplanted labeled cells were monitored. For simplicity, four-cell recipient embryos are depicted, although morula-stage embryos were used as both donors and recipients. The results, summarized in the graphs, show that outer cells overwhelmingly form trophectoderm and that inner cells tend to become part of the ICM, but also form considerable trophectoderm.
[Part (b) R. L. Gardner & J. Nichols, “An investigation of the fate of cells transplanted orthotopically between morulae/nascent blastocysts in the mouse,” 1991, Human Reproduction6(1):25–35, by permission of Oxford University Press.]
The fate of a cell in the early embryo—TE or ICM—is determined by the cell’s location. If a labeled cell is placed on the outside of a very early embryo, it is likely to form extraembryonic tissues, while a cell placed inside an embryo is likely to form embryonic tissues (Figure 21-4b, c). Gene expression measurements of each stage of early development show dramatic changes in which genes are expressed. Even these very early embryos use Wnt, Notch, and TGF-β signals to regulate gene expression (see Chapter 16).
Both ICM and TE cells are stem cells: each starts its own distinct lineage and divides prolifically to produce diverse populations of cells. It is the ICM stem cells that we turn our attention to in the next section.