Sequential Opening and Closing of Voltage-Gated Na+ and K+ Channels Generate Action Potentials
The cycle of changes in membrane potential and return to the resting value that constitutes an action potential lasts 1–2 milliseconds and can occur hundreds of times a second in a typical neuron (see Figure 22-2). These cyclical changes in the membrane potential result first from the opening and closing of a number of voltage-gated Na+channels (that is, channels opened by a change in membrane potential) in a segment of the axonal plasma membrane, and then from the opening and closing of voltage-gated K+channels. The role of these channels in the generation of action potentials was elucidated in classic studies done on the giant axon of the squid, in which multiple microelectrodes can be inserted without causing damage to the integrity of the plasma membrane. However, the same basic mechanism is used by all neurons.
Voltage-Gated Na+ Channels As just discussed, voltage-gated Na+ channels are closed in resting neurons. A small depolarization of the membrane (as occurs when neurotransmitter stimulates a postsynaptic cell) increases the likelihood that any one channel will open; the greater the depolarization, the greater the probability that a channel will open. Depolarization causes a conformational change in these channel proteins that opens a gate on the cytosolic surface of the pore, permitting Na+ ions to pass through the pore into the cell. Thus the greater the initial membrane depolarization, the more voltage-gated Na+ channels that open and the more Na+ ions that enter.
As Na+ ions flow inward through opened channels, the excess positive charges on the cytosolic face and negative charges on the exoplasmic face diffuse a short distance away from the initial site of depolarization. This passive spread of positive charges on the cytosolic face and negative charges on the external face depolarizes (makes the inside less negative) adjacent segments of the plasma membrane, causing opening of additional voltage-gated Na+ channels in these segments and an increase in Na+ influx. As more Na+ ions enter the cell, the inside of the cell membrane becomes more depolarized, causing the opening of yet more voltage-gated Na+ channels and even more membrane depolarization, setting into motion an explosive entry of Na+ ions. For a fraction of a millisecond, the permeability of this small segment of the membrane to Na+ becomes vastly greater than that for K+, and the membrane potential approaches ENa, the equilibrium potential for a membrane permeable only to Na+ ions. As the membrane potential approaches ENa, however, further net inward movement of Na+ ions ceases, since the concentration gradient of Na+ ions (outside > inside) is now offset by the inside-positive membrane potential. The action potential is, at its peak, close to the value of ENa.
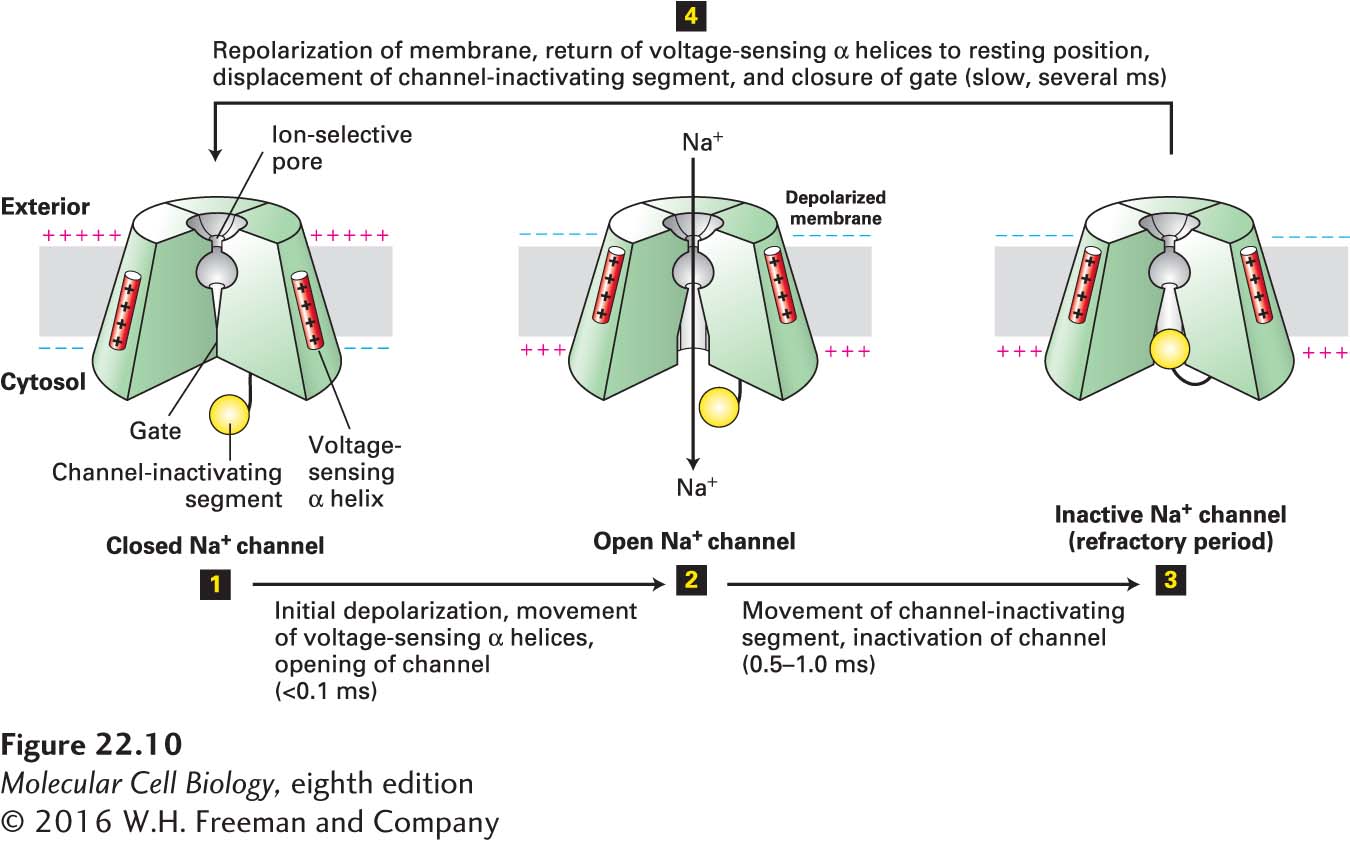
Figure 22-10 schematically depicts the critical structural features of voltage-gated Na+ channels and the conformational changes that cause their opening and closing. In the resting state, a segment of the protein on the cytosolic face—the gate—obstructs the central pore, preventing passage of ions. The channel contains four positively charged voltage-sensing α helices; in the resting state these helices are attracted to the inside-negative surface of the plasma membrane. A small depolarization of the membrane triggers movement of these voltage-sensing helices toward the negative charges that are building up on the exoplasmic surface, causing a conformational change in the gate that opens the channel and allows Na+ ion flow. After about 1 ms, further Na+ influx is prevented by movement of the cytosol-facing channel-inactivating segment into the open channel, blocking any further movement of Na+ ions. As long as the membrane remains depolarized, the channel-inactivating segment remains in the channel opening; during this refractory period, the channel is inactivated and cannot be reopened. A few milliseconds after the inside-negative resting potential is reestablished, the channel-inactivating segment swings away from the pore and the voltage-sensing α helices return to their resting position near the cytosolic surface of the membrane. Thus the channel returns to the closed resting state, once again able to be opened by depolarization. Note the important distinction between “closed” channels and those that are “inactive” as depicted in Figure 22-10.
FIGURE 22-10Operational model of the voltage-gated Na+channel. As in the K+ channel depicted in Figure 11-20, four transmembrane domains in the protein contribute to the central pore through which ions move. The critical components that control movement of Na+ ions are shown here in the cutaway views depicting three of the four transmembrane domains. In the closed, resting state, the voltage-sensing α helices, which have positively charged side chains every third residue, are attracted to the negative charges on the cytosolic side of the resting membrane. This keeps the gate segment near the cytosolic face in a “closed” position that blocks the channel, preventing entry of Na+ ions (step 1). In response to a small depolarization, the voltage-sensing helices move through the phospholipid bilayer toward the outer membrane surface, causing an immediate conformational change in the gate at the cytosolic face of the protein that opens the channel (step 2). Within a fraction of a millisecond the channel-inactivating segment moves into the open channel, preventing passage of further ions (step 3). Once the membrane is repolarized, the voltage-sensing helices return to the resting position, the channel-inactivating segment is displaced from the channel opening, and the gate closes; the protein reverts to the closed, resting state and can be opened again by depolarization (step 4). See W. A. Catterall, 2001, Nature409:988; and S. B. Long et al., 2007, Nature450:376.
Page 1036
Voltage-Gated K+ Channels The repolarization of the membrane that occurs during the refractory period is due largely to opening of voltage-gated K+ channels. The subsequent increased efflux of K+ from the cytosol removes the excess positive charges from the cytosolic face of the plasma membrane (i.e., makes it more negative), thereby restoring the inside-negative resting potential. For a brief instant, the membrane actually becomes hyperpolarized; at the peak of this hyperpolarization, the potential approaches EK, which is more negative than the resting potential (see Figure 22-2).
Opening of the voltage-gated K+ channels is induced by the large depolarization of the action potential. Unlike voltage-gated Na+ channels, most types of voltage-gated K+ channels remain open as long as the membrane is depolarized, and close only when the membrane potential has returned to an inside-negative value. Because the voltage-gated K+ channels open slightly after the initial depolarization, at the height of the action potential, they sometimes are called delayed K+channels. Eventually all the voltage-gated K+ and Na+ channels return to their closed resting states. The only open channels in this baseline condition are the nongated K+ channels that generate the resting membrane potential, which soon returns to its usual value of −60 to −70 mV (see Figure 22-9a).
While the flow of Na+ and K+ ions alters membrane potential dramatically as it is depolarized, hyperpolarized, and repolarized during an action potential cycle, it is important to note that the exchange of these ions across the membrane is small compared to the overall numbers of Na+ and K+ ions in the cytosol and extracellular space. Thus the conduction of action potentials in neurons does not directly require the Na+/K+ pumps that maintain their ion concentration gradients, as we will see shortly.
Page 1037
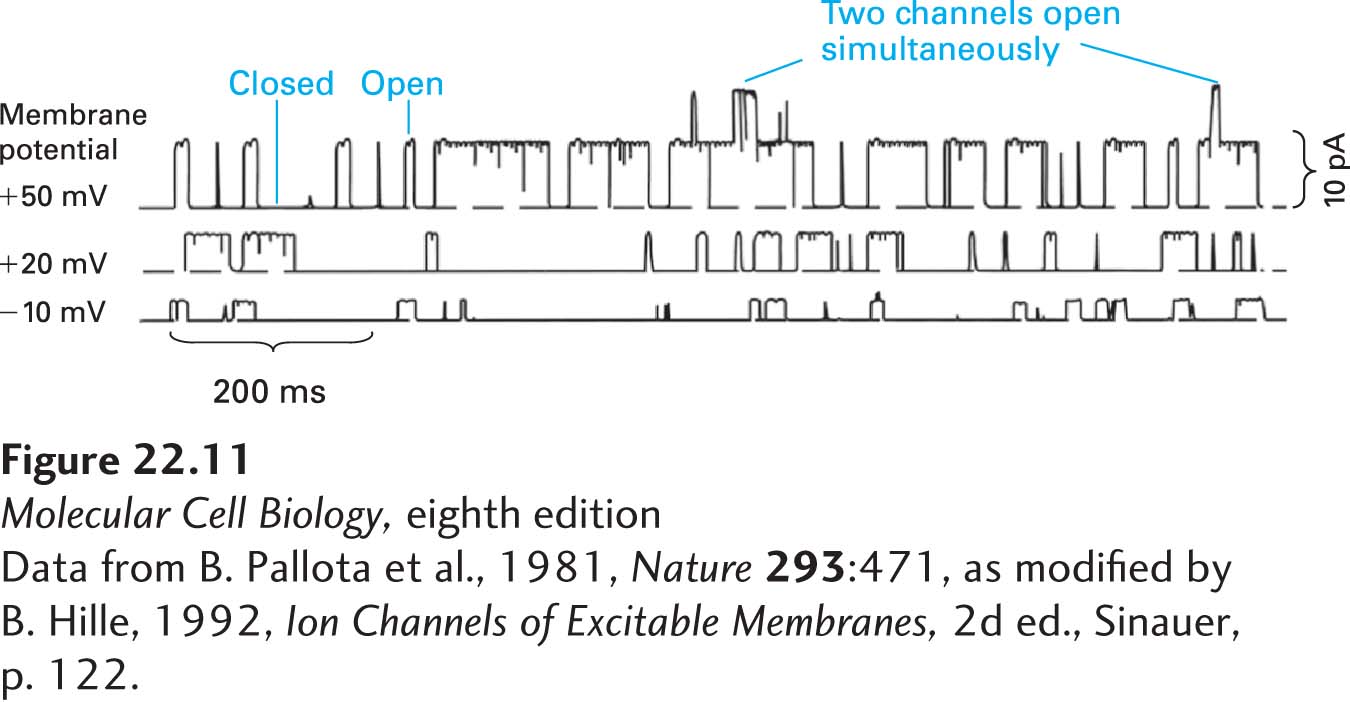
Voltage-gated Na+ channels are difficult to study using patch-clamp techniques, but the patch-clamp tracings in Figure 22-11 reveal the essential properties of voltage-gated K+ channels (see Figure 11-22 for a description of patch clamping). In this experiment, small segments of a neuronal plasma membrane were held clamped at different voltages, and the flux of electric charges through the patch due to flow of K+ ions through open K+ channels was measured. At the modest depolarizing voltage of −10 mV, the channels in the membrane patch open infrequently and remain open for only a few milliseconds, as judged, respectively, by the number and width of the upward blips on the tracings. Further, the ion flux through them is rather small, as measured by the electric current passing through each open channel (the height of the blips). Depolarizing the membrane further to +20 mV causes these channels to open about twice as frequently; also, more K+ ions move through each open channel (the height of the blips is greater) because the force driving cytosolic K+ ions outward is greater at a membrane potential of +20 mV than at −10 mV. Depolarizing the membrane further to +50 mV, the value at the peak of an action potential, causes opening of more K+ channels and also increases the flux of K+ through them. Thus by opening during the peak of the action potential, these K+ channels permit the outward movement of K+ ions and repolarization of the membrane potential while the voltage-gated Na+ channels are being closed and inactivated.
[Data from B. Pallota et al., 1981, Nature293:471, as modified by B. Hille, 1992, Ion Channels of Excitable Membranes, 2d ed., Sinauer, p. 122.]
EXPERIMENTAL FIGURE 22-11Probability of channel opening and current flux through individual voltage-gated K+channels increases with the extent of membrane depolarization. These patch-clamp tracings were obtained from patches of neuronal plasma membrane clamped at three different potentials, +50, +20, and –10 mV. The upward deviations in the current indicate the opening of K+ channels and movement of K+ ions outward (cytosolic to exoplasmic face) across the membrane. Increasing the membrane depolarization (i.e., the clamping voltage) from –10 mV to +50 mV increases the probability a channel will open, the time it stays open, and the amount of electric current (numbers of ions) that passes through it. pA = picoamperes.
[Data from B. Pallota et al., 1981, Nature293:471, as modified by B. Hille, 1992, Ion Channels of Excitable Membranes, 2d ed., Sinauer, p. 122.]
More than 100 voltage-gated K+ channel proteins have been identified in humans and other vertebrates. As we discuss later, all these channel proteins have a similar overall structure, but they exhibit different voltage dependencies, conductivities, channel kinetics, and other functional properties. Many open only at strongly depolarizing voltages, a property required for generation of the maximal depolarization characteristic of the action potential before repolarization of the membrane begins.
Given the fundamental role voltage-gated Na+ and K+ channels play in determining action potential firing, it is not surprising that mutations in these channels give rise to inherited, monogenic human epilepsies. Epilepsies are seizure disorders that affect about 1% of the population and that result from excessive synchronized neuronal activity in the brain. While epilepsy can arise from a variety of causes, including abnormal brain development, brain injury, and drug and alcohol abuse, some forms of epilepsy are caused by mutations in genes that encode ion channels. These diseases are called channelopathies. Human genetic studies have identified specific mutations in the Nav1.1 voltage-gated Na+ channel that cause generalized epilepsy with febrile seizures, while mutations in the Kv7.2 and Kv7.3 voltage-gated K+ channels cause another form of epilepsy called benign familial neonatal convulsions. Mutations in voltage-gated Na+ and K+ channels cause neuronal hyperexcitability in a variety of ways, including by altering Na+ channel inactivation or blocking K+ channel–dependent repolarization of neurons, both of which prolong the duration of action potentials, or by lowering the threshold for triggering action potentials, for example, by decreasing the ratio of inhibitory to excitatory inputs onto neurons.