Movement of the Channel-Inactivating Segment into the Open Pore Blocks Ion Flow
An important characteristic of most voltage-gated channels is inactivation; that is, soon after opening they close spontaneously, forming an inactive channel that will not reopen until the membrane is repolarized. In the resting state, the globular balls at the N-termini of the four subunits in a voltage-gated K+ channel are free in the cytosol (see Figure 22-13). Several milliseconds after the channel is opened by depolarization, one ball moves through an opening between two of the subunits and binds in a hydrophobic pocket in the pore’s central cavity, blocking the flow of K+ ions (see Figure 22-10). After a few milliseconds, the ball is displaced from the pore, and the protein reverts to the closed, resting state. The ball-and-chain domains in K+ channels are functionally equivalent to the channel-inactivating segment in Na+ channels.
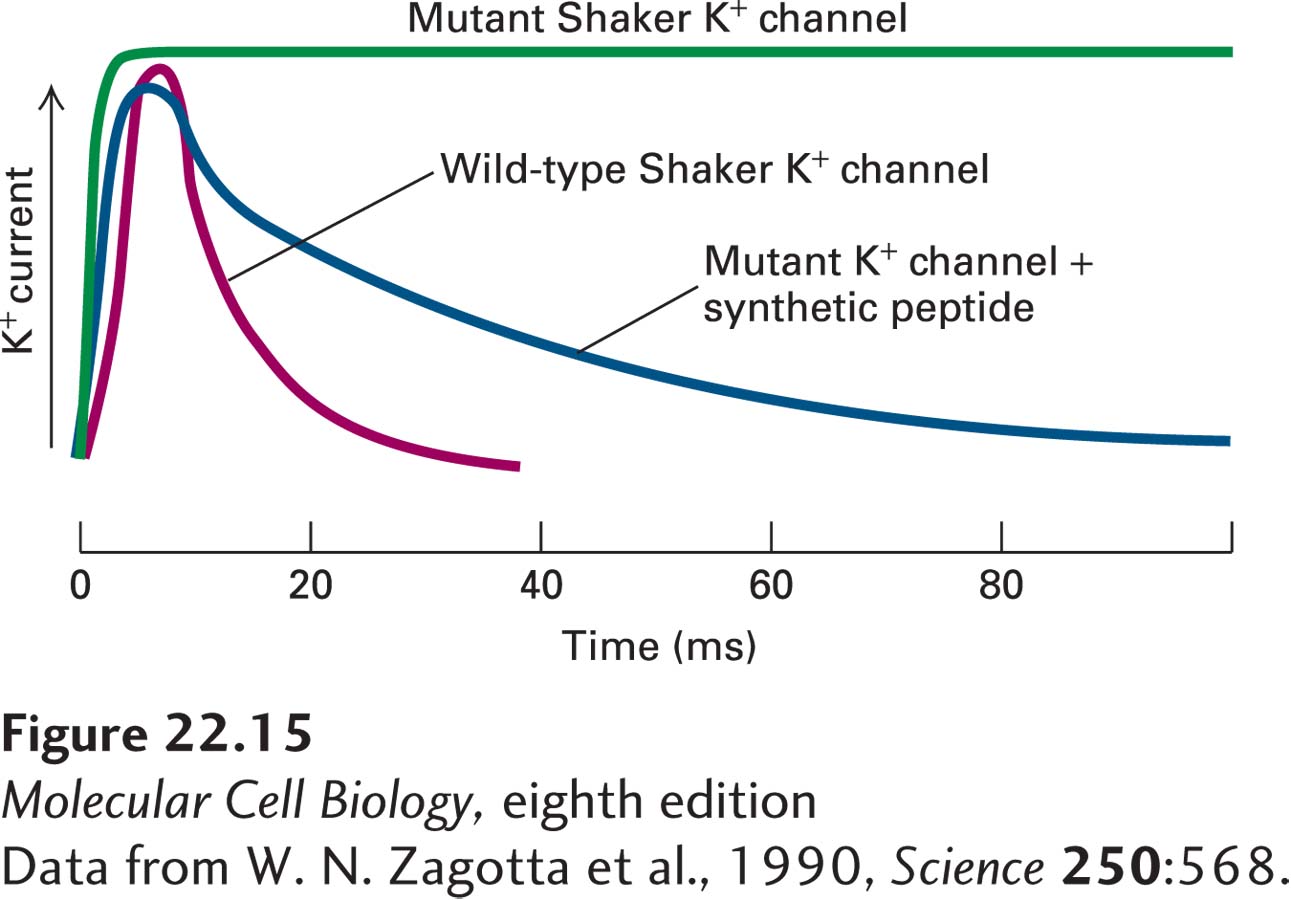
The experimental results shown in Figure 22-15 demonstrate that inactivation of K+ channels depends on the ball domains, occurs after channel opening, and does not require the ball domains to be covalently linked to the channel protein. In other experiments, mutant K+ channels lacking portions of the ~40-residue chain connecting the ball to the S1 helix were expressed in frog oocytes. Patch clamp measurements of channel activity showed that the shorter the chain, the more rapid the inactivation, as if a ball attached to a shorter chain can move into the open channel more readily. Conversely, addition of random amino acids to lengthen the normal chain slows channel inactivation.
[Data from W. N. Zagotta et al., 1990, Science250:568.]
EXPERIMENTAL FIGURE 22-15Experiments with a mutant K+channel lacking the N-terminal globular domains support the ball-and-chain inactivation model. The wild-type Shaker K+ channel and a mutant form lacking the amino acids composing the N-terminal ball were expressed in Xenopus oocytes. The activity of the channels was monitored by the patch-clamp technique. When patches were depolarized from 0 to +30 mV, the wild-type channel opened for ~5 ms and then closed (red curve). The mutant channel opened normally, but could not close (green curve). When a chemically synthesized ball peptide was added to the cytosolic face of the patch, the mutant channel opened normally and then closed (blue curve). This demonstrated that the added peptide inactivated the channel after it opened and that the ball does not have to be tethered to the protein in order to function.
[Data from W. N. Zagotta et al., 1990, Science250:568.]
Page 1043
The single channel-inactivating segment in voltage-gated Na+ channels contains a conserved hydrophobic motif composed of isoleucine, phenylalanine, methionine, and threonine (see Figure 22-13b). Like the longer ball-and-chain domain in K+ channels, this segment folds into and blocks the Na+-conducting pore until the membrane is repolarized.
The technique of molecular dynamics has provided further insights into the structure and function of voltage-gated ion channels. Molecular dynamics involves computer simulations of the physical movement of molecules and atoms in time, basing the simulations on experimental data derived from structural, biochemical, and molecular studies of the molecule of interest. Molecular dynamic studies of a prokaryotic voltage-gated Na+ channel provide a cinematic view of voltage sensing, pore opening, and gate inactivation.