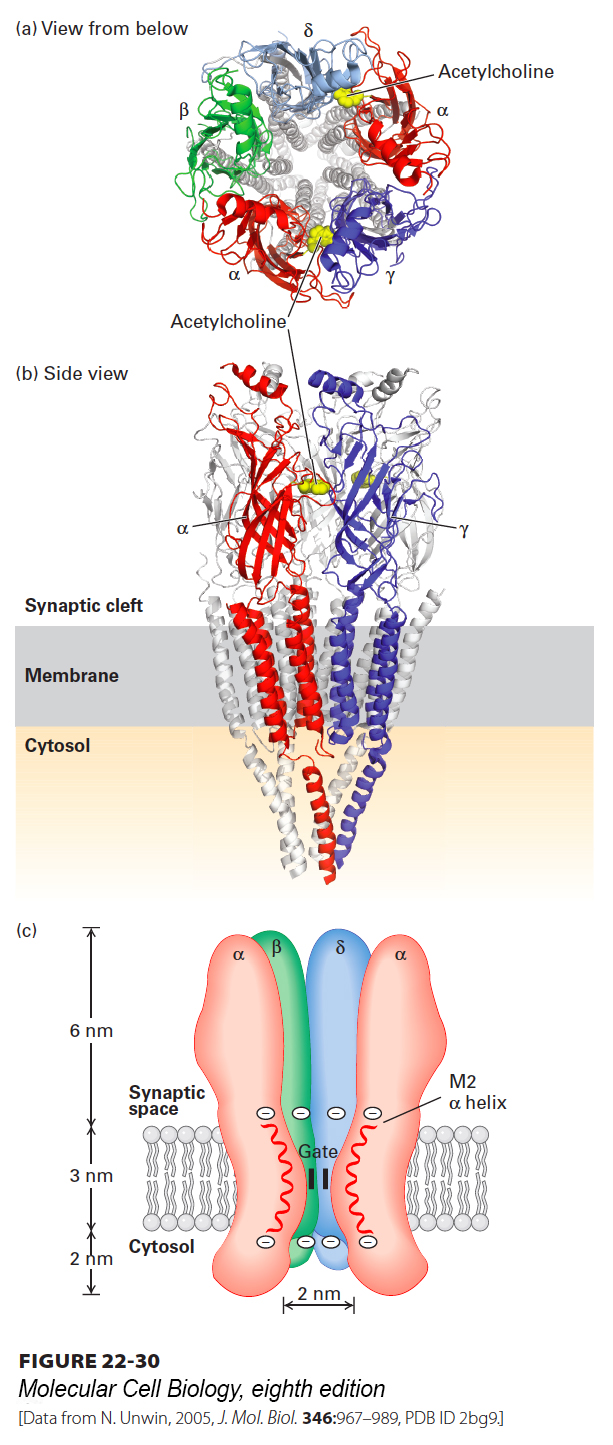
All Five Subunits in the Nicotinic Acetylcholine Receptor Contribute to the Ion Channel
The excitatory nicotinic acetylcholine receptor, found at many nerve-muscle synapses, was the first ligand-gated ion channel to be purified, cloned, and characterized at the molecular level, and provides a paradigm for other neurotransmitter-gated ion channels. The acetylcholine receptor from skeletal muscle is a pentameric protein with a subunit composition of α2βγδ. These four different subunit types have considerable sequence homology with each other; on average, about 35–40 percent of the residues in any two subunits are similar, suggesting that they all derived from a common ancestral gene. The complete receptor has five-fold symmetry, and the actual cation channel is a tapered central pore lined by homologous segments from each of the five subunits (Figure 22-30). The channel opens when the receptor cooperatively binds two acetylcholine molecules to sites located at the interfaces of the αδ and αγ subunits, as shown in Figure 22-30a. Once acetylcholine is bound to a receptor, the channel is opened within a few microseconds. Studies measuring the receptor’s permeability to different small cations suggest that the open ion channel is, at its narrowest, about 0.65–0.80 nm in diameter, in agreement with estimates from electron micrographs. This would be sufficient to allow passage of both Na+ and K+ ions with their shell of bound water molecules.
[Data from N. Unwin, 2005, J. Mol. Biol.346:967–989, PDB ID 2bg9.]
FIGURE 22-30Three-dimensional structure of the nicotinic acetylcholine receptor. Three-dimensional molecular structure of the Torpedo nicotinic acetylcholine receptor as viewed (a) from the synaptic cleft and (b) parallel to the plane of the membrane. For clarity, only the front two subunits, α and γ, are highlighted in (b) (colors: α, red; β, green; γ, blue; δ, light blue). The two acetylcholine-binding sites are located about 3 nm from the membrane surface and are highlighted in yellow; only the one at the α γ interface is shown in panel (b). (c) Schematic cutaway model of the pentameric receptor in the membrane. Each subunit has four membrane-spanning α helices, M1–M4; the M2 α helix (red) faces the central pore. Aspartate and glutamate side chains form two rings of negative charges, one at each end of the M2 helices, that help exclude anions from and attract cations to the channel. The gate, which is opened by binding of acetylcholine, lies within the pore.
[Data from N. Unwin, 2005, J. Mol. Biol.346:967–989, PDB ID 2bg9.]
We have discussed the neuromuscular junction as an excellent example of how neurotransmitters and their receptors work. Like acetylcholine, glutamate, a principal neurotransmitter in the vertebrate brain, uses two main types of receptors. One class, termed ionotropic glutamate receptors, are ligand-gated channels that allow the flow of K+, Na+, and sometimes Ca2+ in response to glutamate binding and that work along the same principles as AChR. Glutamate also binds to a second class of receptors, coupled to G proteins. Later in this chapter we will see how such G protein–coupled receptors (GPCRs) and ion channels function as receptors for odorants and tastants that activate various sensory nerve cells. To cover all of the neurotransmitter receptors, ion channels, and other signaling proteins that function in the brain would require a book much larger than this one!