Animals as diverse as snails and humans sense noxious events (the process termed nociception); pain receptors, called nociceptors, respond to mechanical change, heat, and certain toxic chemicals. Pain serves to alert us to events such as tissue damage that are capable of producing injury and evokes behaviors that promote tissue healing. Persistent pain in response to tissue injury is common, and many individuals suffer from chronic pain. Thus understanding both acute and chronic pain is a major research goal, as is the development of new types of drugs to treat pain.
One of the first mammalian pain receptors to be cloned and identified was TRPV1, a Na+/Ca2+ channel that is found in many sensory pain neurons of the peripheral nervous system and is activated by a wide variety of exogenous and endogenous physical and chemical stimuli. The best-known activators of TRPV1 are heat greater than 43 °C, acidic pH, and capsaicin, the molecule that makes chili peppers seem hot. Activation of TRPV1 receptors leads to painful, burning sensations. Numerous TRPV1 antagonists have been developed by pharmaceutical companies as possible pain medications. However, a major side effect that has limited the utility of these drugs is that they result in an elevation in body temperature; this suggests that one “normal” function of TRPV1 is to sense and regulate body temperature, and that the drugs inhibit this function.
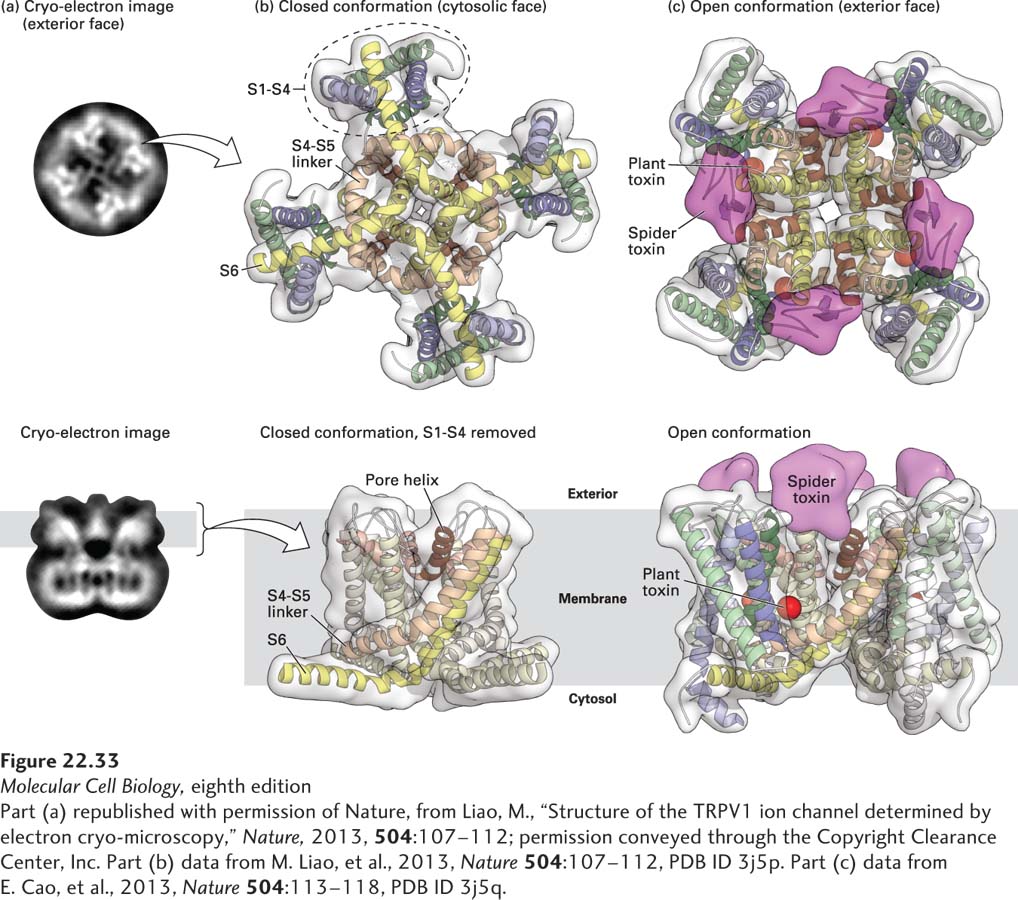
In a recent landmark study, scientists used single-particle cryoelectron microscopy (cryoEM, see Chapter 3) to obtain a high-resolution (0.34 nm) model of the rat TRPV1 channel in the closed configuration and in two open configurations, one bound to capsaicin and the other bound to two potent TRPV1 activators, one from plant and the other from spider venom. As shown in Figure 22-33, these studies revealed that the TRPV1 channel structure is similar to that of voltage-gated ion channels (see Figure 22-13), composed of four symmetrical subunits with six transmembrane helices (S1–S6) each. However, the charged amino acids in S1–S4 that function as voltage sensors in voltage-gated ion channels are replaced by aromatic residues in TRPV1. This stabilizes the channel core so that instead of moving like voltage sensors upon depolarization, the TRPV S1–S4 helices provide an anchor for movements within the pore that are triggered by ligand binding. Two constrictions, or gates, were identified in the pore region. The spider toxin bound to the extracellular surface of the channel, near the pore helix, and locked open the extracellular end of the channel. Capsaicin and the plant toxin bound to a site deep within the membrane toward the cytoplasmic end of the pore, with binding increasing the diameter of the pore. These findings indicate that the TRPV channel undergoes dual gating.
[Part (a) republished with permission of Nature, from Liao, M., “Structure of the TRPV1 ion channel determined by electron cryo-microscopy,” Nature, 2013, 504:107–112; permission conveyed through the Copyright Clearance Center, Inc. Part (b) data from M. Liao, et al., 2013, Nature504:107–112, PDB ID 3j5p. Part (c) data from E. Cao, et al., 2013, Nature504:113–118, PDB ID 3j5q.]
FIGURE 22-33Single-particle cyroelectron microscopy high-resolution structure of the TRPV1 channel. The high-resolution structure of the rat TRPV1 channel was obtained by single-particle cryoelectron microscopy at 0.34 nm resolution. (a) Photomicrographs of the two-dimensional structure of the tetrameric TRPV1 channel embedded in a thin layer of vitreous ice, with a face view of the channel in the top panel, and a side view in the bottom panel. (b, top) Ribbon diagram of a bottom view of the channel that focuses on the S1–S4 transmembrane domains, and the pore domain formed by S5 and S6, together with linking pore (P) loops. The S1–S4 domain is similar in structure to the voltage-sensing domains in the voltage-gated K+ and Na+ channels (see Figure 22-14), but differ in that they do not move. (b, bottom) Ribbon diagram of the side view of the channel in the closed conformation, focusing on the pore domain that is formed by S5-P-S6. (c) The open conformation was stabilized by incubating the channel with two agonists, a spider toxin (in magenta) and a plant toxin (in red). Cryoelectron density maps reveal that the spider toxin (magenta) binds to external domains of the channel, linking two subunits of the channel together via its two globular cysteine-knot domains, while the plant toxin (red) binds to a region deep within the pore. Capsaicin binds to the same sites as the plant toxin (not shown). Binding of agonists to two distinct sites indicates that the TRPV1 channel is dually gated, allowing for significant modulation of channel function.
[Part (a) republished with permission of Nature, from Liao, M., “Structure of the TRPV1 ion channel determined by electron cryo-microscopy,” Nature, 2013, 504:107–112; permission conveyed through the Copyright Clearance Center, Inc. Part (b) data from M. Liao, et al., 2013, Nature504:107–112, PDB ID 3j5p. Part (c) data from E. Cao, et al., 2013, Nature504:113–118, PDB ID 3j5q.]
Page 1063
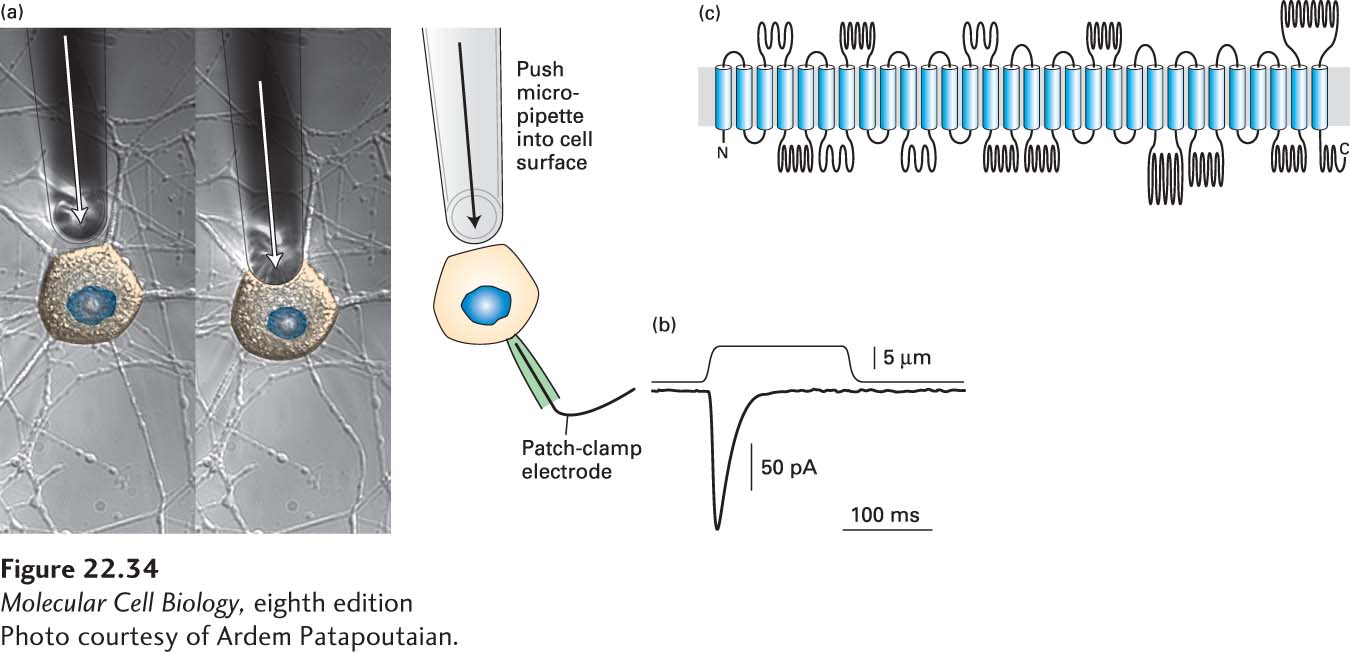
Two channels were discovered in 2010 that directly convert mechanical stimuli into cation conductance in vertebrate cells, called Piezo1 and Piezo2 (from the Greek word piesi, which means “pressure”). Both form large cation-selective channels composed of four identical subunits, with each subunit containing over 30 membrane-spanning domains, creating a channel whose molecular weight is about 1.2 million daltons, and that has between 120 and 160 transmembrane segments! Expression of Piezo1 or Piezo2 induces mechanosensitive-cation currents in these cells. This can be assayed by expressing the channels in cell culture, and using calcium imaging to monitor the response of the cells to stretch induced by poking the cells with a small glass pipette (Figure 22-34). Reduction of Piezo2 expression in dorsal root ganglion sensory neurons in mice reduced their mechanosensitivity, and knockout of the single Piezo homolog in Drosophila melanogaster resulted in flies with severely reduced behavioral responses to noxious mechanical stimuli. Together, these experiments show that Piezo channels mediate mechanical signal transduction.
Page 1064
[Photo courtesy of Ardem Patapoutaian.]
FIGURE 22-34Piezo channels are mechanotransducers. (a) To identify channels that transduced mechanical information, cDNAs encoding transmembrane proteins were expressed in heterologous cells, and the response to mechanical perturbation with a glass pipette was determined by patch clamp recording (shown) or by calcium imaging (not shown). (b) When Piezo1 or Piezo2 cDNAs are expressed in cultured cells, poking the cell with a glass pipette elicits a strong inward current. (c) Piezo1 and 2 form homotetrameric cation channels. Each subunit is extremely large, with over 2000 amino acid residues and over 30 transmembrane domains. The assembled channel thus contains over 120 membrane spanning domains, and has a mass of over 1.2 million daltons (about the same mass as the small subunit of the ribosome)!