Five Primary Tastes Are Sensed by Subsets of Cells in Each Taste Bud
We taste many chemicals, all of which are hydrophilic and nonvolatile molecules floating in saliva. All tastes are sensed on all areas of the tongue, and selective cells respond preferentially to certain tastes. Like the other senses, that of taste likely evolved to increase an animal’s chance of survival. Many toxic substances taste bitter or acidic, and nourishing foods are broken down into molecules that taste sweet (e.g., sugars), salty, or umami (e.g., the meaty or savory taste of monosodium glutamate and other amino acids). Animals (including humans) can never be certain exactly what enters their mouth; the sense of taste enables an animal to make a quick decision—eat it, or get rid of it. Taste is less demanding of the nervous system than olfaction, because fewer types of molecules are monitored. What is impressive is the sensitivity of taste; bitter molecules can be detected at concentrations as low as 10−12 M.
There are receptors for salty, sweet, sour, umami, and bitter tastes in all parts of the tongue. The receptors are of two different types: channel proteins for salty and sour tastes and seven-transmembrane-domain proteins (G protein–coupled receptors) for sweetness, umami, and bitterness. Specific membrane receptors that detect fatty acids are present on taste bud cells, and fatty taste may come to be recognized as a sixth basic taste quality.
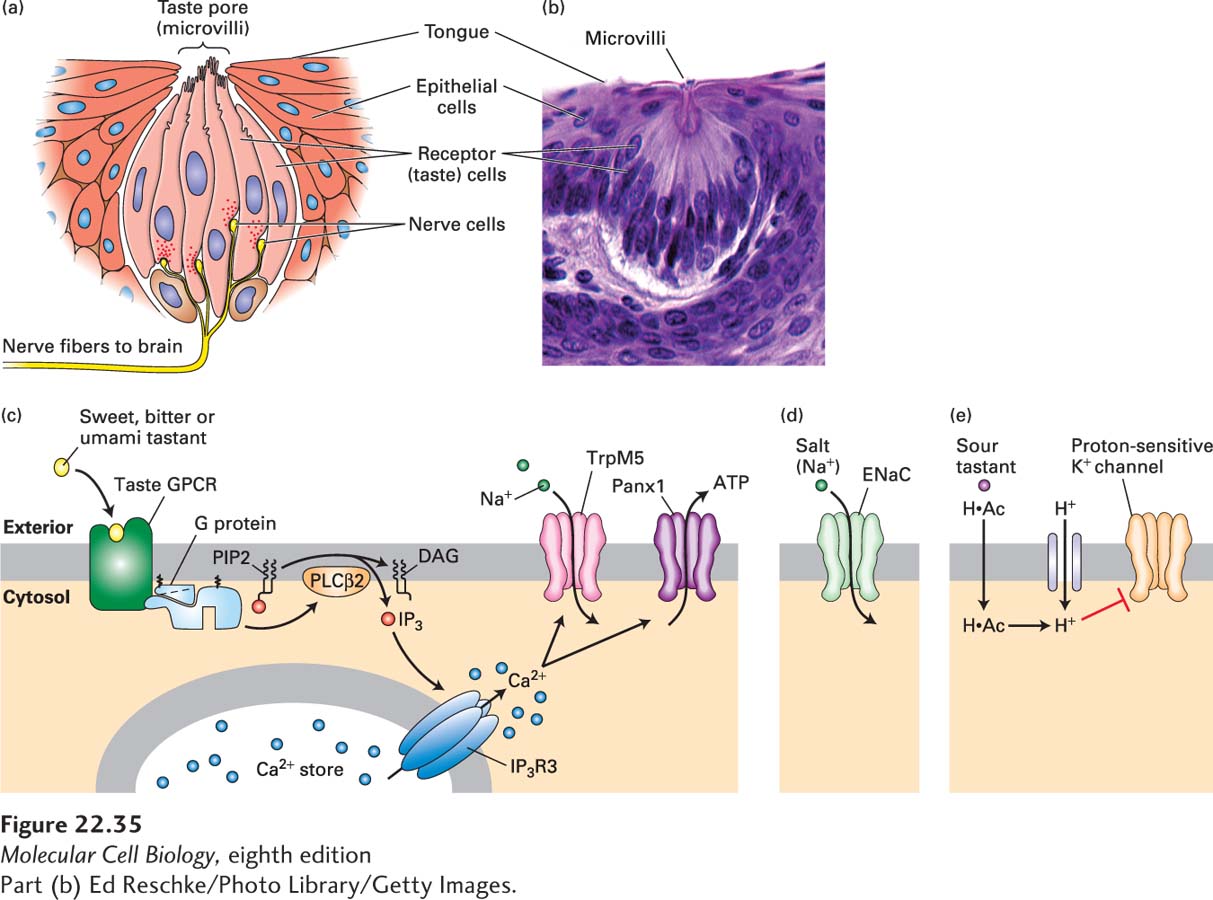
Taste buds are located in bumps in the tongue called papillae; each bud has a pore through which fluid carries solutes inside. Each taste bud has about 50–100 taste cells (Figure 22-35a, b), which are epithelial cells but with some of the functions of neurons. Microvilli on the taste cells’ apical tips bear the taste receptors, directly contacting the external environment in the oral cavity and thus experiencing wide fluctuations in food-derived molecules as well as the presence of potentially harmful compounds. Cells in the tongue and other parts of the mouth are subjected to a lot of wear and tear, and taste bud cells are continuously replaced by cell divisions in the underlying epithelium. (A taste bud cell in a rat has a lifetime of 10 days.)
[Part (b) Ed Reschke/Photo Library/Getty Images.]
FIGURE 22-35The sense of taste. (a) The taste cells (pink) in a mammalian taste bud contact the nerve cells (yellow). The chemical signals arrive at the microvilli seen at the top. (b) Micrograph of a mammalian taste bud, showing the receptor cells. The microvilli are barely visible at the top of the taste bud, indicated by the label. (c) Sweet, bitter, and umami ligands bind specific taste GPCRs expressed in Type II receptor cells, activating a phosphoinositide pathway that elevates cytosolic Ca2+ · Ca2+ in turn binds to and opens a Ca2+-gated Na+ channel, TrpM5, leading to an influx of Na+ and membrane depolarization. The combined action of elevated Ca2+ and membrane depolarization opens the large pores of an unusual membrane channel termed Panx1, resulting in release of ATP and probably other signaling molecules into the extracellular space. ATP and these other molecules stimulate the nerve cells that will ultimately carry the information to the brain. (d) Salt is detected by direct permeation of Na+ ions through membrane ion channels, including the ENaC channel, directly depolarizing the plasma membrane. (e) Organic acids like acetic acid diffuse in their protonated form (H·Ac) through the plasma membrane and dissociate into an anion and proton, acidifying the cytosol. Entry of strong acids like HCl is facilitated by a proton channel in the apical membrane of the sour-sensing cells that enables protons to reach the cytosol. Intracellular H+ is believed to block a proton-sensitive K+ channel (as yet unidentified) and thus depolarize the membrane. Voltage-gated Ca2+ channels would open, leading to an elevation in cytosolic Ca2+ that triggers exocytosis of synaptic vesicles that are not depicted. See N. Chaudhari and S. D. Roper, 2010, J. Cell Biol.190:285; and S. Frings, 2010, PNAS107:21955.
[Part (b) Ed Reschke/Photo Library/Getty Images.]
Reception of a taste signal causes cell depolarization that triggers action potentials; these in turn cause Ca2+ uptake through voltage-dependent Ca2+ channels and release of neurotransmitters (Figure 22-35c–e). Taste cells do not grow axons; instead, they signal over short distances to adjacent neurons. These neurons convey the information about taste through multiple connections to a region of the cortex that is specialized for taste, called the insula. To determine how the insula knows that a salty taste receptor as compared to a sweet taste receptor has been activated, scientists performed two-photon imaging (Chapter 4) of insula in mice after presentation of a specific tastant. They used calcium indicators to detect the neurons that were activated, and in this way were able to monitor the activation of large numbers of neurons as calcium-dependent increases in fluorescence. These experiments revealed that four of the tastes—sweet, bitter, umami and salty—are represented in separate, nonoverlapping regions within the insula, thereby demonstrating the existence of a gustotopic map in the brain that mediates our representation of taste.
Page 1065
Bitter Taste Bitter tastants are diverse and are detected by a family of about 25–30 different G protein–coupled receptors (GPCRs) known as T2Rs. As depicted in Figure 22-35c, all of these GPCRs activate a particular Gα isoform, called gustducin, which is expressed only in taste cells. However, it is the released ubiquitous Gβγ subunit of the heterotrimeric G protein that binds to and activates a specific isoform of phospholipase Cβ, which in turn generates IP3. IP3 triggers Ca2+ release from the endoplasmic reticulum (see Figure 15-34). Ca2+ in turn binds to and opens a Ca2+-gated Na+ channel, TRPM5, a member of the TRP family of ion channels, leading to an influx of Na+ and membrane depolarization. The combined action of elevated Ca2+ and membrane depolarization opens the large pores of a membrane channel termed Panx1, resulting in release of ATP and probably other signaling molecules into the extracellular space. ATP is then thought to stimulate the nerve cells that will ultimately carry the taste information to the brain.
Page 1066
Different bitter taste molecules are quite distinct in structure, which probably accounts for the need for the diverse family of T2Rs. Some T2Rs bind only 2–4 bitter-tasting compounds, whereas others bind a wider variety of bitter compounds. The first member of the T2R family to be identified came from human genetics studies that showed an important bitterness-detection gene on chromosome 5. Mice that have five amino acid changes in the T2R protein T2R5 are unable to detect the bitter taste of cycloheximide (a protein synthesis inhibitor; see Table 4-1). Multiple T2R types are often expressed in the same taste cell, and about 15 percent of all taste cells express T2Rs.
A dramatic gene regulation swap experiment was done to demonstrate the role of T2R proteins. Mice were engineered to express a bitter-taste receptor, a T2R protein, in cells that normally detect sweet tastants that attract mice. The mice developed a strong attraction for bitter tastes, evidently because the cells continued to send a “go and eat this” signal even though they were detecting bitter tastants. This experiment demonstrates that the specificity of taste cells is determined within the cells themselves, and that the signals they send are interpreted according to the neural connections made by that class of cells. It implies that the T2R-expressing sweet taste receptors were wired to the region of the insula that receives “sweet” signals, and that it was thus represented as a sweet taste.
Sweet and Umami Tastes Sweet and umami tastants are detected by a GPCR family called the T1Rs, which are related to the T2Rs and that also transduce signals through a phosphoinositide signaling pathway. The three mammalian T1Rs differ from one another in a small number of amino acids. The T1Rs have very large extracellular domains that comprise the taste-binding domain of the protein. In the taste-sensing glutamate receptor, the extracellular domain closes around glutamate in a way that is described as analogous to a Venus flytrap. Unlike most GPCRs, which generally function as monomers, T1Rs form homodimers and heterodimers, which is thought to increase the repertoire of molecules that can act as signals. However, the code of responses to different molecules is still under investigation. Mice lacking T1R2 or T1R3 fail to detect sugar; it is thought that the actual receptor is a heterodimer of the two. T1R3 appears to be a receptor for both sweet tastes and umami, and that is because it detects sweets when combined with T1R2 and umami when it combines with T1R1. Accordingly, taste cells express T1R1 or T1R2 but not both, as otherwise they would send an ambiguous message to the brain.
Interestingly, sweet-taste receptors are also found on the surface of certain endocrine cells in the gut; these cells also express gustducin and several other taste transduction proteins. The presence of glucose in the gut causes these cells to secrete the hormone glucagon-like peptide-1 (GLP-1), which in turn regulates appetite, and enhances insulin secretion and gut motility. Thus certain cells of the gut “taste” glucose through the same mechanisms used by taste cells of the tongue.
Salty Taste The taste of salt is elicited by a wide range of Na+ concentrations, from 10 mM to 500 mM. Salt is sensed by a member of a family of Na+ channels called ENaC channels (Figure 22-35d). Indeed, knocking out a critical ENaC subunit in taste buds impaired salty-taste detection in mice. The influx of Na+ through the channel depolarizes the taste cell, leading to neurotransmitter release. The role of ENaC channels as salt sensors is evolutionarily ancient; ENaC proteins also detect salt when expressed in insects. In Drosophila, taste sensors are located in multiple places including the legs, so when the fly steps on something tasty, the proboscis extends to explore it further.
Sour Taste Perception of sourness is due to the detection of H+ ions. Many sour tastants are weak organic acids (e.g., acetic acid in vinegar), which in their protonated forms diffuse through the plasma membrane. They then dissociate into an anion and a proton, which acidifies the cytosol. Strong acids like HCl are detected by a proton channel in the apical membrane of the sour-sensing cells that enables protons to reach the cytosol. Regardless of how the intracellular H+ concentration is increased, protons are believed to block an as-yet-unidentified proton-sensitive K+ channel in mammals and thus depolarize the membrane (Figure 22-35e). As with salt detection, voltage-gated Ca2+ channels would then open, elevate cytosolic Ca2+, and thus trigger exocytosis of neurotransmitter-filled synaptic vesicles.