The perception of volatile airborne chemicals imposes different demands than the perception of light, sound, touch, or taste. Light is sensed by only four rhodopsin molecules, tuned to different wavelengths. Sound is detected by mechanical effects through hairs that are tuned to different wavelengths. Touch and pain requires a small number of different gated ion channels. The sense of taste measures a small number of substances dissolved in water. In contrast to all these other senses, olfactory systems can discriminate between many hundreds of volatile molecules moving through air. Discrimination between a large number of chemicals is useful in finding food or a mate, sensing pheromones, and avoiding predators, toxins, and fires. Olfactory receptors work with enormous sensitivity. Male moths, for example, can detect single molecules of the signals sent drifting through the air by females. In order to cope with so many signals, the olfactory system employs a large family of olfactory receptor proteins. Humans have about 700 olfactory receptor genes, of which about half are functional (the rest are unproductive pseudogenes), a remarkably large proportion of the estimated 20,000 human genes. Mice are more efficient, with more than 1200 olfactory receptor genes, of which about 800 are functional. That means 3 percent of the mouse genome is composed of olfactory receptor genes. Drosophila has about 60 olfactory receptor genes. In this section we will examine how olfactory receptor genes are employed, and how the brain can recognize which odor has been sensed—the initial stages of interpretation of our chemical world. Odor molecules are called odorants. They have diverse chemical structures, so olfactory receptors face some of the same challenges faced by antibodies and hormone receptors—the need to bind and distinguish many variants of relatively small molecules.
Page 1067
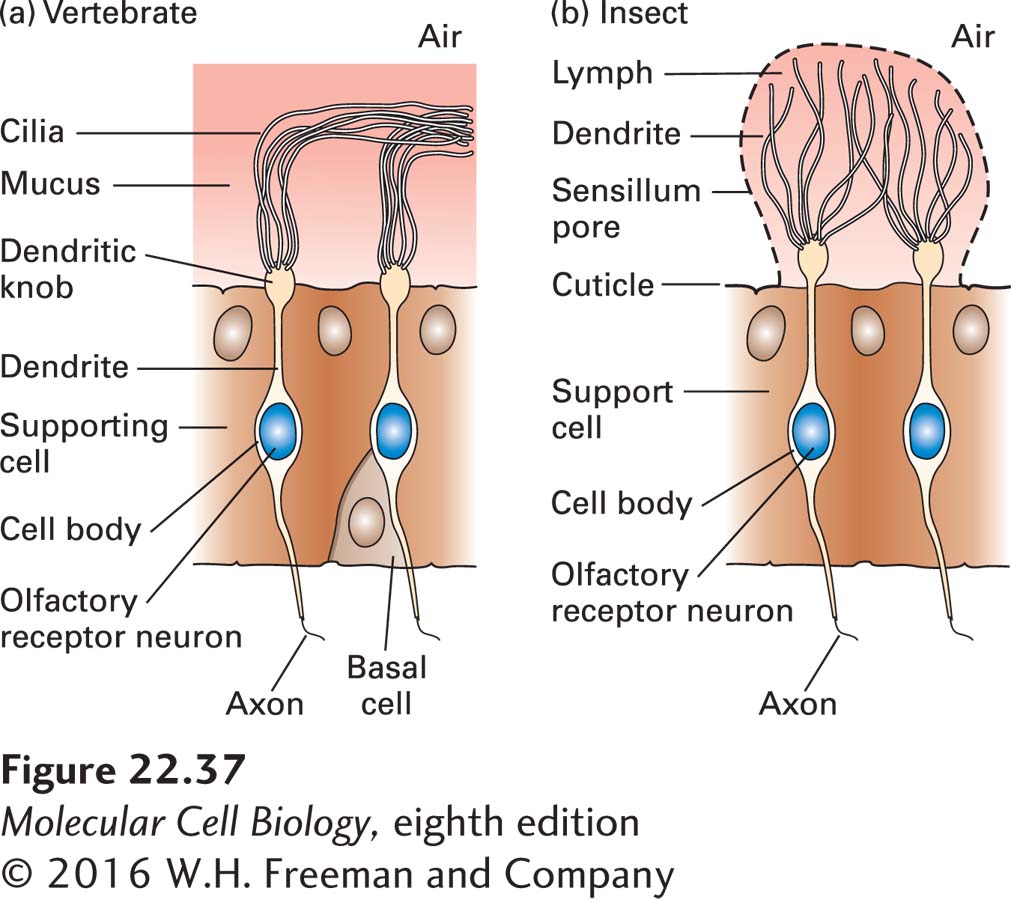
Olfactory receptors are seven-transmembrane-domain proteins (Figure 22-36). In mammals, olfactory receptors are produced by cells of the nasal epithelium. These cells, called olfactory receptor neurons (ORNs), transduce the chemical signal into action potentials. Each ORN extends a single dendrite to the luminal surface of the epithelium, from which immotile cilia extend to bind inhaled odorants from the air (Figure 22-37a). These olfactory sensory cilia are enriched in the odorant receptors and signal transduction proteins that mediate the initial transduction events. In Drosophila, ORNs have similar structures and are located in the antennae (Figure 22-37b).
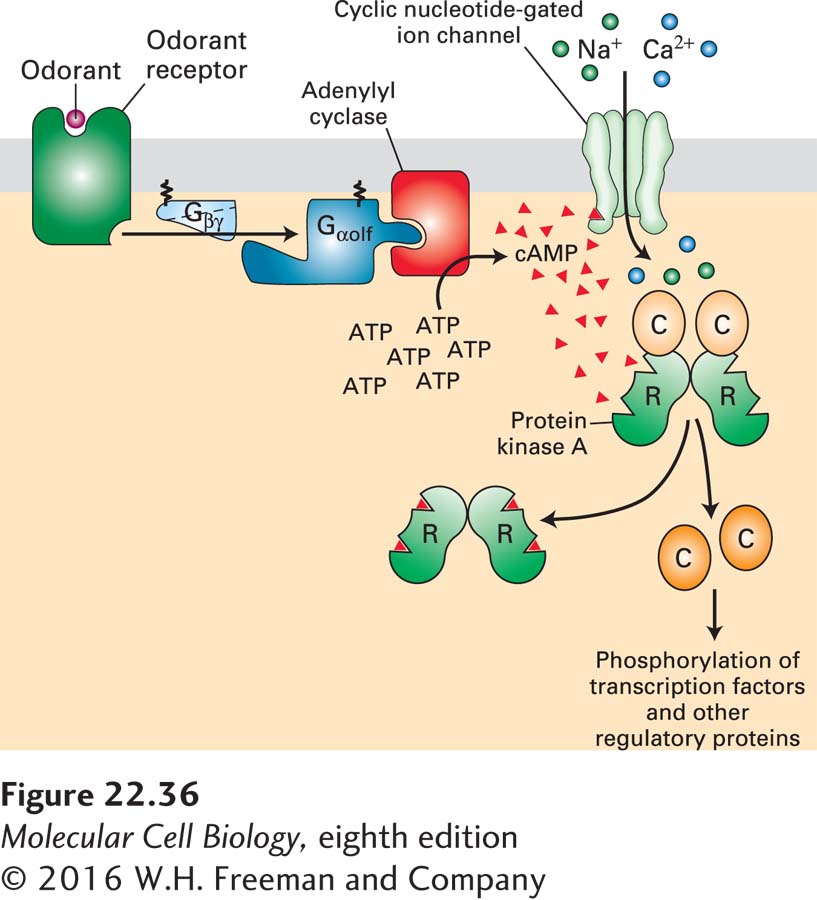
FIGURE 22-36Signal transduction from the olfactory GPCRs. Binding of an odorant to its cognate odorant receptor (OR) triggers activation of the trimeric G protein Gαolf · Gβγ, releasing the active Gαolf · GTP. Activated Gαolf · GTP in turn activates type III adenylyl cyclase (AC3), leading to the production of cyclic AMP (cAMP) from ATP. Molecules of cAMP bind to and open the cyclic nucleotide–gated (CNG) ion channel, leading to the influx of Na+ and Ca2+ and depolarizing the cell. cAMP also activates protein kinase A (PKA), which phosphorylates and thus regulates transcription factors and other intracellular proteins.
FIGURE 22-37Structures of olfactory receptor neurons. Across a vast span of evolutionary distance—vertebrate and insect—olfactory receptor neurons have similar forms. (a) Vertebrate olfactory receptor neurons have one dendrite, which ends in a dendritic knob; from each dendritic knob, approximately 15 cilia extend into the nasal mucus. (b) Insect olfactory receptor neurons are morphologically similar: the bipolar neuron gives rise to a single basal axon that projects to an olfactory glomerulus in the antennal lobe. At its apical side it has a single dendritic process, from which sensory cilia extend. See U. B. Kaupp, 2010, Nature Rev. Neurosci. 11:188–200.
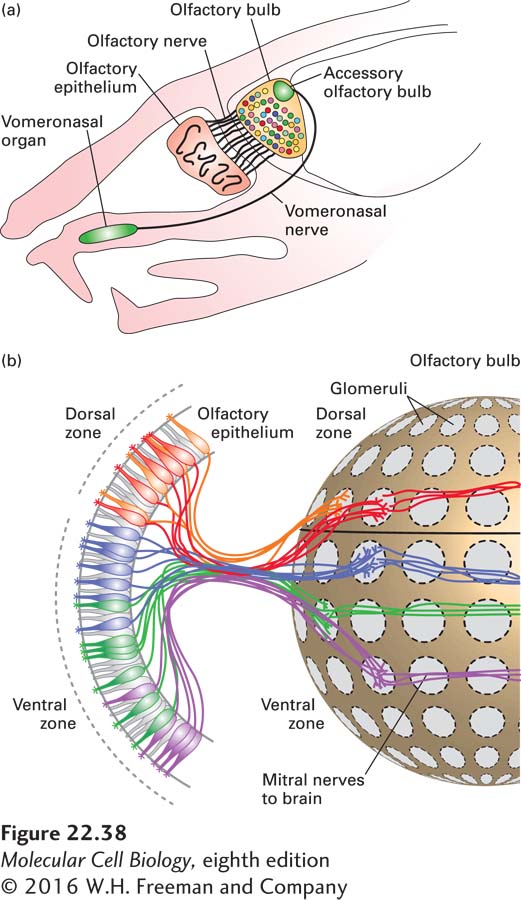
In both mammals and Drosophila the ORNs project their axons to the next higher level of the nervous system, which in mammals is located in the olfactory bulb of the brain. The ORN axons synapse with dendrites from mitral neurons in mammals (called projection neurons in insects); these synapses occur in the clusters of synaptic structures called glomeruli. The mitral neurons connect to higher olfactory centers in the brain (Figure 22-38).
FIGURE 22-38The anatomy of olfaction in the mouse. (a) Schematic representation of a sagittal section through an adult mouse head. Axons of the olfactory receptor neurons (ORNs) in the main olfactory epithelium bundle to form the olfactory nerve and innervate the olfactory bulb. Each ORN of the main olfactory epithelium expresses only one odorant receptor gene. The vomeronasal organ and the accessory olfactory bulb are involved in pheromone sensing. (b) All of the olfactory receptor neurons that express a single type of receptor send their axons to the same glomerulus. In this figure each color represents the neural connections for each distinct expressed receptor. The glomeruli are located in the olfactory bulb near the brain; in the glomeruli, the ORNs synapse with mitral neurons; each mitral neuron has its dendrites localized to a single glomerulus and its corresponding ORNs, thus carrying information about a particular odorant to higher centers of the brain. Each glomerulus thus receives innervation from sensory neurons expressing a single odorant receptor, providing the anatomical basis of the olfactory sensory map. See T. Komiyama and l. Luo, 2005, Curr. Opin. Neurobiol.16:67–73 and S. Demaria and J. Ngai, 2010, J. Cell Biol.191:443.
Humans vary markedly in their ability to detect certain odors. Some cannot detect the steroid androstenone, a compound derived from testosterone and found in human sweat. Some describe the odor as pleasant and musky, while others compare it to the smell of dirty socks. These differences are all ascribed to inactivating missense mutations in the gene encoding the single androstenone GPCR. Individuals with two copies of the wild-type allele perceive androstenone as unpleasant, whereas those possessing one or no functional alleles perceive androstenone as less unpleasant or undetectable.
Despite the vast number of olfactory receptors, all generate the same intracellular signals through activation of the same trimeric G protein: Gαolf · Gβγ (see Figure 22-36). Gαolf is expressed mainly in olfactory neurons. Like Gαs, the active Gαolf · GTP formed after ligand binding activates an adenylyl cyclase that leads to the production of cyclic AMP (cAMP; see Figure 15-25). Two downstream signaling pathways are activated by cAMP. It binds to a site on the cytosolic face of a cyclic nucleotide–gated (CNG) Na+/Ca2+ channel, opening the channel and leading to an influx of Na+ and Ca2+and local depolarization of the cell membrane. This odorant-induced depolarization in the olfactory dendrites spreads throughout the neuronal membrane, resulting in opening of voltage-gated Na+ channels in the axon hillock and the generation of action potentials. Molecules of cAMP also activate protein kinase A (PKA), which phosphorylates and thus regulates transcription factors and other intracellular proteins.