Neural Stem Cells Form Nerve and Glial Cells in the Central Nervous System
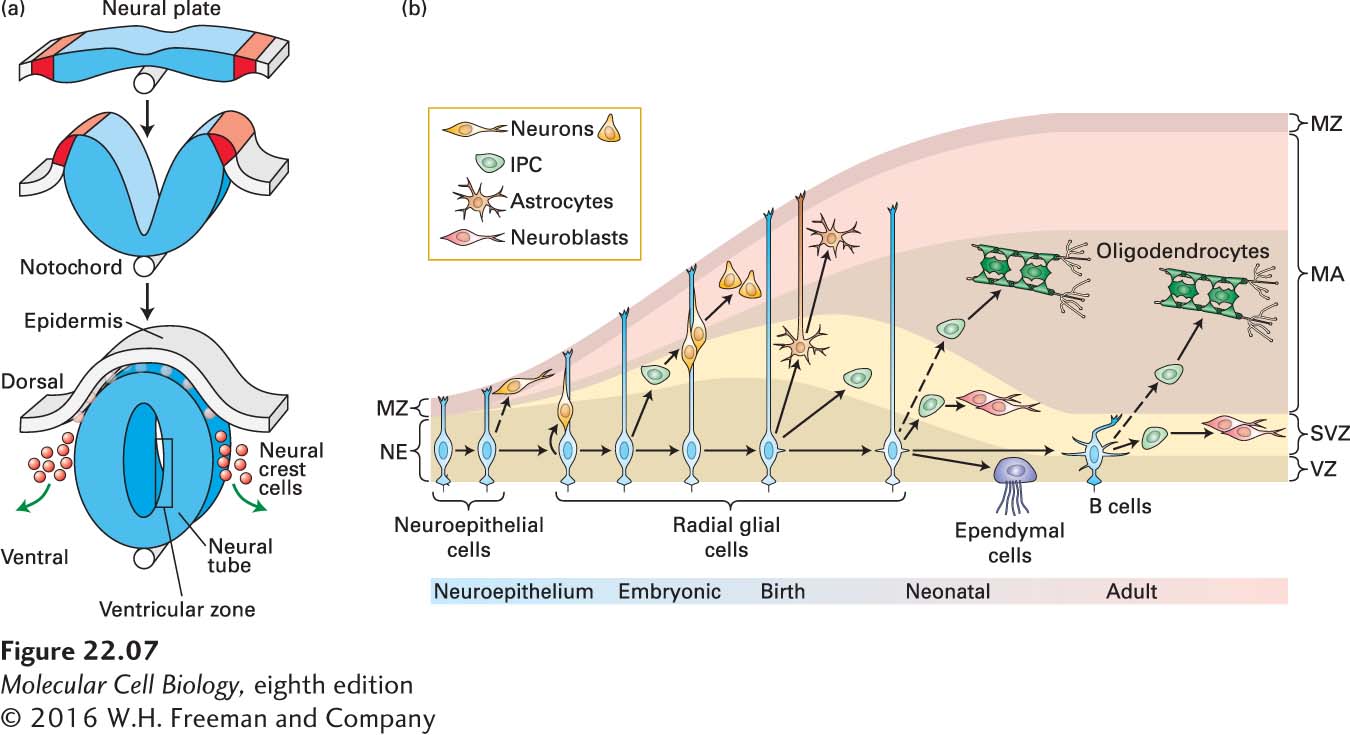
The great interest in the formation of the nervous system and in finding better ways to prevent or treat neurodegenerative diseases through cell replacement therapies has made the characterization of neural stem cells, and their differentiation into mature neurons and glia, an important goal. Much of what we understand about neural and glial stem cells comes from studies of embryonic brain development. The earliest stages of vertebrate neural development involve the rolling up of a tube of ectoderm (the cell layer that lines the outside of the embryo) that extends the length of the embryo from head to tail (Figure 22-7a). This neural tube will form the brain and spinal cord. Initially the thickness of the tube is a single layer of cells, and these cells, referred to as neuroepithelial cells, serve as the embryonic neural stem cells (NSCs) that will give rise to the entire central nervous system. The inside of the neural tube will expand in the forebrain to form the fluid-filled compartments called ventricles, and the cellular layer lining the neural tube, where most cell division takes place, is called the ventricular zone (VZ).
FIGURE 22-7Formation of the neural tube and division of neural stem cells. (a) Early in vertebrate development a part of the ectoderm rolls up and separates from the rest of the cells. This forms the epidermis (gray) and the neural tube (blue). Near the interface between the two, neural crest cells form and then migrate to contribute to skin pigmentation, nerve formation, craniofacial skeleton, heart valves, peripheral neurons, and other structures. The notochord, a rod of mesoderm for which chordates are named, provides signals that affect cell fates in the neural tube. The interior of the neural tube will become a fluid-filled series of chambers called ventricles. Neural stem cells located adjacent to the ventricles, described as being in the ventricular zone (VZ), will divide to form neurons that migrate radially outward to form the layers of the nervous system. (b) Early in development, in the neuroepithelium (NE), neuroepithelial cells divide symmetrically to generate more neuroepithelial cells. Some are also thought to generate early neurons. As development progresses and the brain epithelium thickens, neuroepithelial cells convert into radial glial cells (RGCs). RGCs divide symmetrically or asymmetrically to generate neurons or intermediate progenitor cells (IPCs), which in turn generate neurons. RGCs continue to elongate, and send an apical process down to the VZ and a basal process up to contact the meninges. Near the end of embryonic development, RGCs detach from the NE and convert into astrocytes, and also generate oligodendrocytes from IPCs. After birth, in neonates, RGCs continue to divide into neurons and oligodendrocytes, through IPCs. Others convert into ependymal cells or into adult SVZ astrocytes called type B cells that function as neural stem or progenitor cells in the subventricular zone (SVZ) of the adult brain. IPC, intermediate progenitor cell; MA, mantle; MZ, marginal zone; NE, neuroepithelium; RG, radial glia; SVZ, subventricular zone; VZ, ventricular zone. See A. Kriegstein and A. Alvarez-Buylla, 2009, Annu. Rev Neurosci.32:149–184.
Labeling and tracing experiments in mouse have been done to determine how cells are born and where they go after birth. The embryonic neuroepithelial cells (NECs), the neural stem and progenitor cells that line the ventricle, can divide symmetrically, producing two daughter stem or progenitor cells side by side (Figure 22-7b), thereby expanding the progenitor population. Around the same time that neuron production begins, the NECs transform into radial glial cells, which are the primary precursor cells during embryonic neurogenesis. Radial glial cells also divide symmetrically into two daughter radial glial cells or asymmetrically, into either another radial glial cell and a differentiated neuron, or a radial glial cell and an intermediate progenitor cell. The intermediate precursor cells move into a region just adjacent to the VZ called the subventricular zone (SVZ), and they in turn give rise to differentiated neurons. Newborn neurons use the radial glia as scaffolds as they migrate away from the VZ toward the surface of the brain, migrating radially outward. In the cerebral cortex, the migrating neurons form successive layers in an inside-out fashion. Later in development, RGCs also give rise to glia, including both astrocytes and oligodendrocytes (Figure 22-7b).
Page 1032
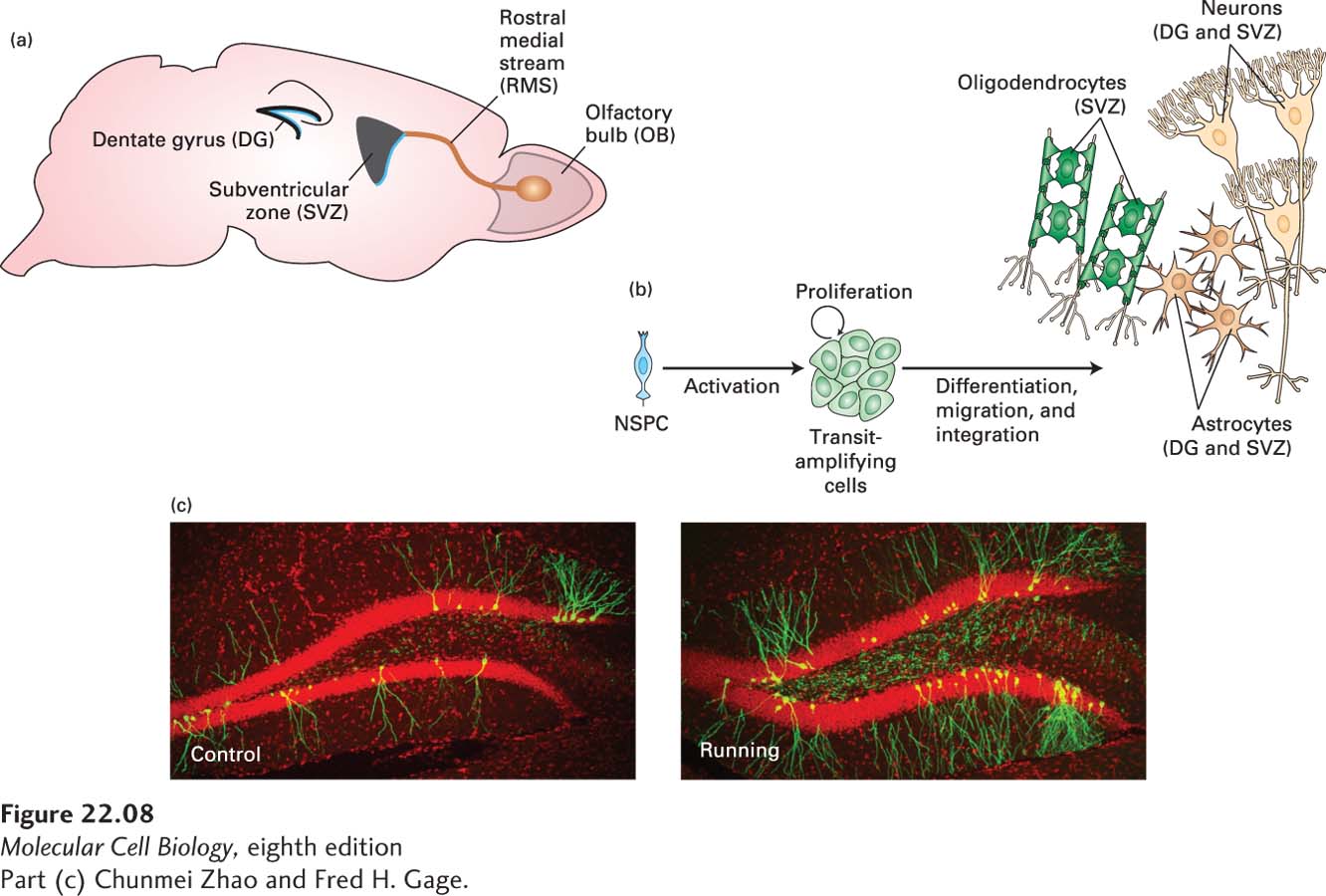
For many years it was believed that no new nerve cells are formed in the adult. Most mammalian brain cells indeed stop dividing by adulthood, but some cells in the lateral ventricle, in a region called the adult SVZ, and in a region of the hippocampus continue to act as stem cells to generate new neurons (Figure 22-8a). Similar to other types of stem cells, these neural stem cells are functionally defined by their ability to self-renew and differentiate into neural lineages, including neurons, astrocytes, and oligodendrocytes (Figure 22-8b). To identify and characterize neural stem cells, cells isolated from the SVZ were cultured with growth factors such as FGF2 or EGF. Some of the cells survived and proliferated in an undifferentiated state; that is, they could self-renew. In the presence of other growth factors, these undifferentiated cells gave rise to neurons, astrocytes, or oligodendrocytes. The successful establishment of self-renewing and multipotent cells from the adult brain provides strong evidence for the presence of nerve stem-cell populations. While the function of these new neurons in the adult brain is not yet understood, studies in rodents have shown that their survival is increased by enriched environments and by exercise (Figure 22-8c).
[Part (c) Chunmei Zhao and Fred H. Gage.]
FIGURE 22-8Neurogenesis in the adult brain. (a) New neurons are born in two regions of the adult brain, the dentate gyrus (DG) in the hippocampus and the subventricular zone (SVZ). Neurons derived from neural stem cells in the svz migrate to the olfactory bulb (OB) via the rostral medial stream (RMS) in mice. (b) Neural stem and precursor cells (NPSCs) can be activated to divide into a class of intermediate precursor cells called transit-amplifying cells, which in turn can divide into astrocytes or neurons in the DG or into astrocytes, neurons, or oligodendrocytes in the SVZ. (c) Newly born neurons in the dentate gyrus were labeled with a retrovirus that expresses GFP. Shown are sections of the dentate gyrus from control mice and from mice that were allowed to exercise on a running wheel in their cages for a week. The newly born neurons are green, and the extensive dendritic branches they have developed indicate that they have survived and have become incorporated into the hippocampus. All the other cells in the hippocampus are labeled with a red nuclear marker. The dense red labeling in the dentate gyrus (the sideways V-shaped structure) are the cell bodies of the granule cells. The other red cell bodies represent glial cells and inhibitory neurons. As this image illustrates, the percentage of granule cells in the dentate gyrus that are newly born is very small, and is significantly increased by running.
[Part (c) Chunmei Zhao and Fred H. Gage.]
Some of the NSCs in the SVZ have properties of astrocytes, such as producing glial fibrillary acidic protein (GFAP). But these NSCs can divide asymmetrically to reproduce themselves and to produce intermediate precursor cells called transit-amplifying cells that in turn divide to form neural precursors (neuroblasts). The SVZ niche is created by mostly unknown signals from the ependymal cells that form a layer lining the ventricle, and by endothelial cells that form blood vessels in the vicinity (see Figure 22-8c). The endothelial cells, and the basal lamina they form, are in direct contact with precursor and stem cells and are believed to be essential in forming the neural stem cell niche. Each neural stem cell extends a single cilium through the ependymal cell layer to directly contact the ventricle. The signals that create the niche are not completely characterized, but there is evidence for a blend of factors, including FGFs, BMPs, IGF, VEGF, TGFα, and BDNF (see Chapter 16 for descriptions of these signaling pathways). The BMPs appear to favor astrocyte differentiation over neural differentiation, one example of cell fate determination control that must remain in proper balance.