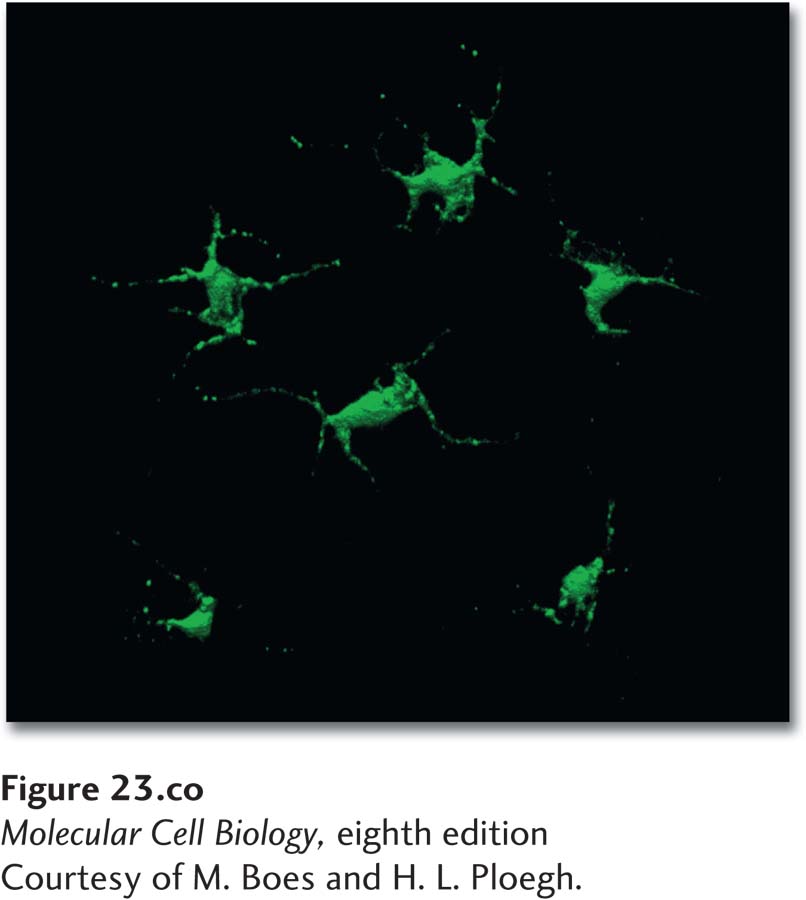
Dendritic cells in the skin have class II MHC molecules on their surface. Those shown here were engineered to express a class II MHC–GFP fusion protein, which fluoresces green.
Immunity is a state of protection against the harmful effects of exposure to pathogens. Host defenses can take many different forms, and all pathogens have found ways to disarm the immune system or manipulate it to their own advantage. Host-pathogen interactions are therefore an evolutionary work in progress. This explains why, despite the evolution of remarkably sophisticated immune systems, pathogenic viruses, bacteria, and parasites continue to pose a threat to human populations. The prevalence of infectious diseases illustrates the imperfections of host defenses. Virtually all pathogens have relatively short generation times compared with the hosts they infect and thus can quickly evolve sophisticated countermeasures against their hosts’ immune system. Seasonal outbreaks of influenza caused by new strains of influenza virus are just one example.
The portion of the immune system that can make adjustments to these threats over time, called the adaptive immune system, changes in response to changing types and abundances of pathogens. Another portion of the immune system, called the innate immune system, serves as the rapid deployment force to deal with invaders. Such sophisticated defenses come at a price: an immune system capable of dealing with a massively diverse collection of rapidly evolving pathogens can sometimes mistake the host’s own tissues for pathogens and mount an attack against its own cells and tissues, a phenomenon called autoimmunity. Even so, we have learned to exploit the workings of the immune system to create vaccines that protect against a variety of infectious diseases. Vaccines are remarkably cost-effective and have contributed to eliminating the scourge of epidemics, such as outbreaks of smallpox.
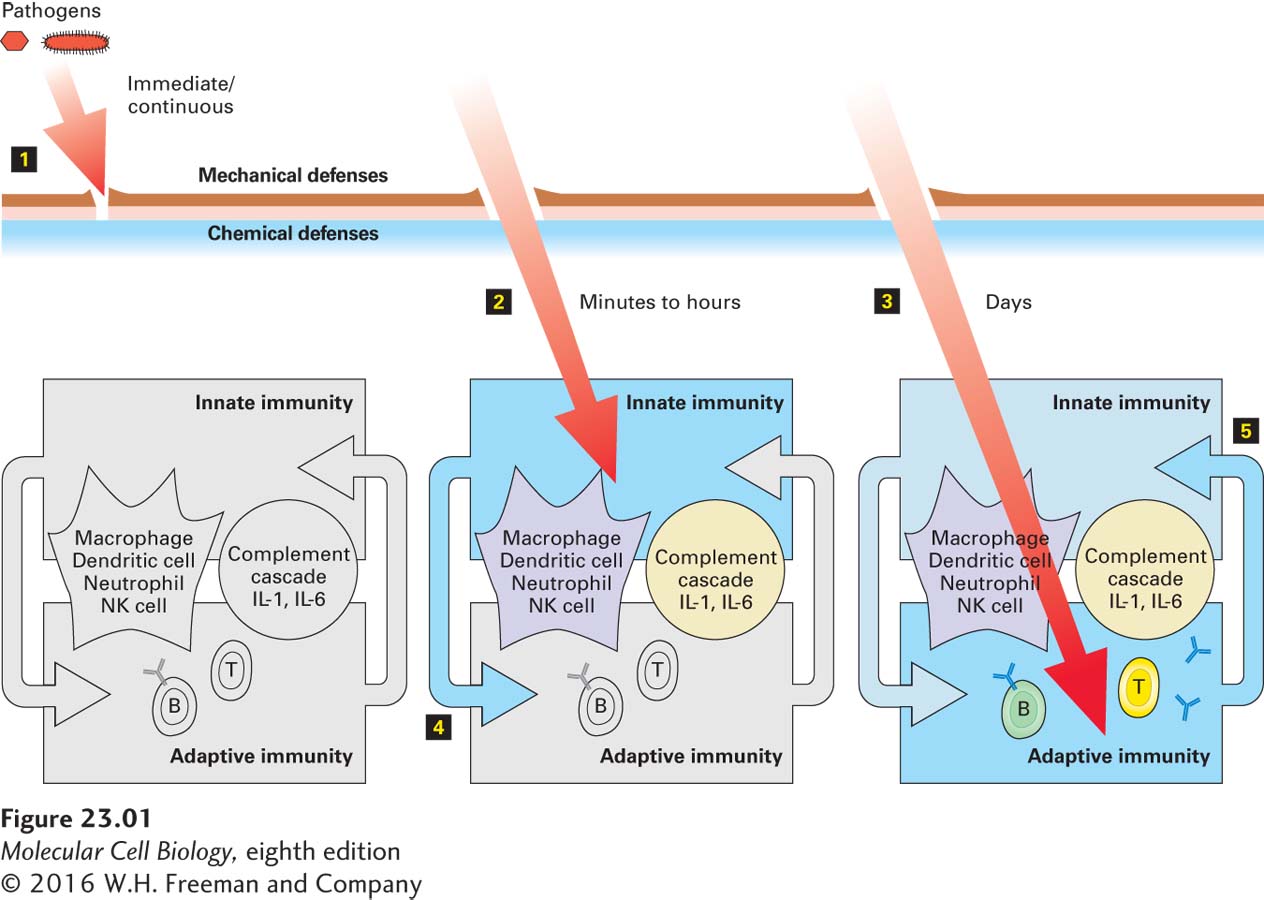
Host defenses comprise three layers: (1) mechanical and chemical defenses, (2) the innate immune system, and (3) the adaptive immune system (Figure 23-1). Mechanical and chemical defenses operate continuously. Innate immune responses, which involve cells and molecules that are present at all times, are rapidly activated (in minutes to hours), but their ability to distinguish among different pathogens is somewhat limited. In contrast, adaptive immune responses take several days to develop fully and are highly specific; that is, they can distinguish between closely related pathogens based on very small molecular differences in their structure.
FIGURE 23-1The three layers of vertebrate immune defenses.Left: Mechanical defenses consist of epithelia and skin. Chemical defenses include the low pH of the gastric environment and antibacterial enzymes in tears. These barriers provide continuous protection against invaders. Pathogens must physically breach these defenses (step 1) to infect the host. Middle: Pathogens that have breached the mechanical and chemical defenses (step 2) are handled by cells and molecules of the innate immune system (blue), which includes phagocytic cells (neutrophils, dendritic cells, macrophages), natural killer (NK) cells, complement proteins, and certain interleukins (IL-1, IL-6). Innate defenses are activated within minutes to hours of infection. Right: Pathogens that are not cleared by the innate immune system are dealt with by the adaptive immune system (step 3), in particular B and T lymphocytes. Full activation of adaptive immunity requires days. The products of an innate response may potentiate an ensuing adaptive response (step 4). Likewise, the products of an adaptive immune response, including antibodies (Y-shaped icons), may enhance innate immunity (step 5). Several cell types and secreted products straddle the fence between the innate and adaptive immune systems and serve to connect these two layers of host defense.
Page 1080
In this chapter, we deal mainly with the vertebrate immune system, with particular emphasis on those molecules, cell types, and pathways that uniquely distinguish the immune system from other types of cells and tissues. Four remarkable features that characterize the vertebrate immune system are specificity, diversity, memory, and tolerance. Specificity is the immune system’s ability to distinguish between closely related substances. Diversity is the system’s capacity to specifically recognize an astoundingly large number (>106) of different molecules. Memory is a host’s ability to recall previously experienced exposure to a foreign substance and more rapidly and effectively defend itself from that substance the next time it is encountered. Tolerance is the ability to avoid mounting an immune-system attack against the host’s own cells and tissues. As we shall see, the immune system achieves specificity and diversity by generating a large number of distinct proteins, such as antibodies and specific cell-surface receptors, each of which can bind very tightly to a target pathogenic molecule, but not to other, perhaps structurally very similar, molecules. Memory and tolerance depend on complex cellular systems we will describe. They are accomplished through the generation of a massively diverse set of cell-surface receptors that bind specific antigens. These receptors have been “trained” to recognize self molecules and are largely unresponsive to self components (self-tolerant).
From a practical perspective, the powers of the immune system can be exploited therapeutically. Today there is a multibillion-dollar market for monoclonal antibodies, which are used in the successful treatment of inflammatory conditions, autoimmune diseases, and cancer. The molecules that constitute the adaptive immune system—antibodies in particular—are also indispensable tools for the cell biologist, as we saw in Chapters 3 and 4. Antibodies allow the visualization and isolation of the molecules they recognize with pinpoint precision. Their ability to do so has been invaluable in the accurate description of the components that make up the cell and its organelles and their localization, both in cells and in tissues. The technique of immunofluorescence, for example, is widely used by cell biologists to study cell morphology and behavior, while immunoblotting (Western blotting) has become an indispensable tool in the study of signal transduction.
Page 1081
Any material that can evoke an immune response is referred to as an antigen. The ways in which these foreign materials are recognized and eliminated involve molecular and cell biological principles unique to the immune system. We begin this chapter with a brief sketch of the organization of the mammalian immune system, introducing the essential players in innate and adaptive immune responses and describing inflammation, a localized response to injury or infection that leads to the activation of immune-system cells and their recruitment to the affected site. In the next two sections, we discuss the structure and function of antibody (or immunoglobulin) molecules, which bind to specific molecular features on antigens, and how variability in antibody structure contributes to the recognition of specific antigens. The enormous diversity of antigens that can be recognized by the adaptive immune system finds its explanation in unique rearrangements of the genetic material in B and T lymphocytes, commonly called B cells and T cells, which are the white blood cells that carry out antigen-specific recognition. These gene rearrangements permit adaptation to a wide variety of pathogens by altering the specificity of antigen-binding receptors on lymphocytes; they also determine cell fate in the course of lymphocyte development.
Although the gene rearrangement mechanisms that give rise to antigen-specific receptors on B and T cells are very similar, the manner in which these receptors bind to (recognize) antigens is very different. The receptors on B cells can interact with intact antigens directly, but the receptors on T cells cannot. Instead, as described in Section 23.4, the receptors on T cells recognize processed forms of antigen cleaved into small peptides, then displayed or “presented” on the surfaces of target cells by specialized cell-surface glycoproteins. These glycoproteins are encoded by genes in a region of the genome called the major histocompatibility complex (MHC). These MHC-encoded glycoproteins, also called MHC products, help determine the host’s ability to mount both T-cell and B-cell responses to antigens.
Understanding these fundamental properties of the immune system has allowed us to answer a number of very practical questions: How can we best make antibodies that afford protection against an infectious agent? How can we raise antibodies to specific proteins we want to study in the laboratory? Knowledge of antigen processing and presentation thus informs both vaccine design to protect against infectious disease and the generation of tools essential for research. MHC-encoded glycoproteins also play a key role in an individual’s development of tolerance for his or her own antigens. We conclude the chapter with an integrated view of the immune response to a pathogen, highlighting the collaboration between different immune-system cells that is required for an effective immune response.