Immunoglobulins Have a Conserved Structure Consisting of Heavy and Light Chains
Immunoglobulins are abundant serum proteins that fall into several classes with distinct structural and functional properties. Immunoglobulins were identified as the class of serum proteins responsible for antibody activity when they were biochemically purified from serum isolated from immunized animals (called antiserum). They were purified based on their abilities to mediate the killing of microbes and to bind directly to their corresponding, or cognate, antigens. Immunoglobulins of the most common class are composed of two identical heavy (H) chains, covalently attached to two identical light (L) chains (Figure 23-9; other classes are described below). The typical immunoglobulin (sometimes abbreviated Ig) therefore has a twofold-symmetric structure, described as H2L2. One H2L2 antibody molecule can usually bind to two antigen molecules (bivalent binding; see below). An exception to this basic H2L2 architecture occurs in the immunoglobulins made by camelids (camels, llamas, vicunas). These animals can make some immunoglobulins that are heavy-chain dimers (H2) and lack light chains.
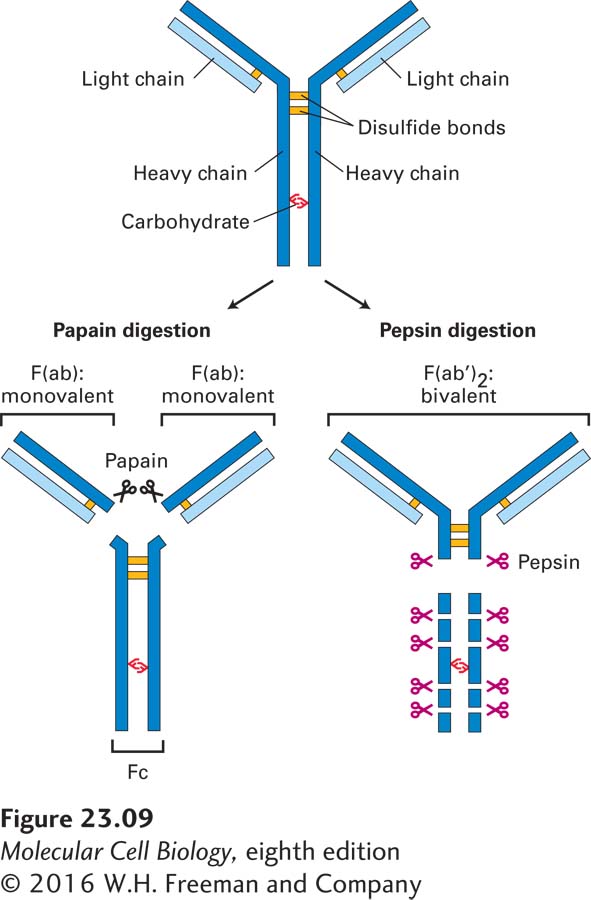
FIGURE 23-9The basic structure of an immunoglobulin molecule. Antibodies are serum proteins also known as immunoglobulins. They are twofold-symmetric structures composed of two identical heavy chains and two identical light chains. Fragmentation of antibodies with proteases yields fragments that retain antigen-binding capacity. The protease papain yields monovalent F(ab) fragments, and the protease pepsin yields bivalent F(ab′)2 fragments. The Fc fragment is unable to bind antigen, but this portion of the intact molecule has other functional properties.
A biochemical approach was used to answer the question of how antibodies manage to distinguish among related molecules—that is, how one antibody can bind to its specific antigen but not to another, structurally very similar, molecule. Proteolytic enzymes were used to fragment immunoglobulins, which are rather large proteins (~150 kDa), to identify the regions in the protein that are directly involved in antigen binding (see Figure 23-9). The protease papain yields fragments, called F(ab) for antigen binding fragment, that can bind a single antigen molecule (monovalent fragments), whereas the protease pepsin yields bivalent fragments, referred to as F(ab′)2 (F = fragment; ab = antibody) that exhibit bivalent binding. These enzymes are commonly used to convert intact immunoglobulin molecules into monovalent or bivalent reagents. Although F(ab) fragments are incapable of cross-linking antigen, F(ab′)2 fragments can do so. Researchers frequently take advantage of this property to cross-link and thus activate surface receptors. Many receptors, such as the EGF receptor, dimerize upon engagement of ligand (ligand-induced dimerization), a prerequisite for full activation of downstream signaling cascades. Many receptors on immune-system cells behave in similar fashion. The portion released upon papain digestion and incapable of antigen binding is called Fc because of its ease of crystallization (F = fragment; c = crystallizable).