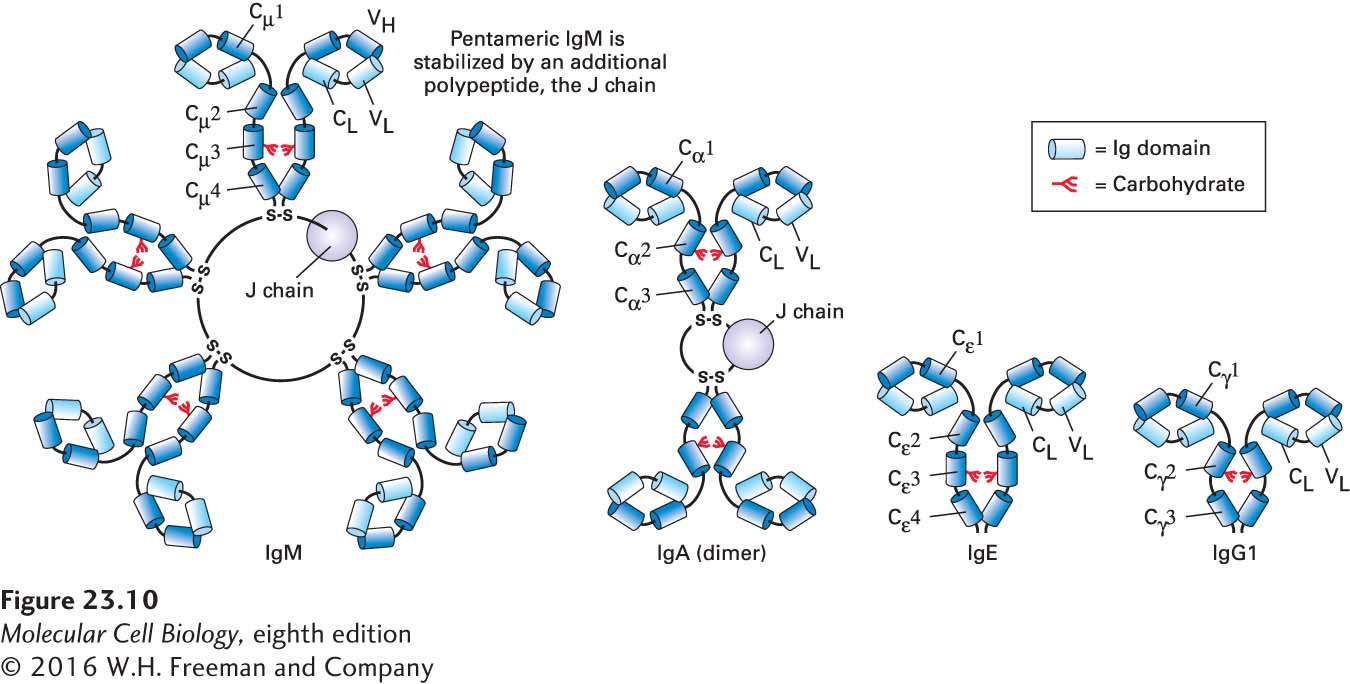
Multiple Immunoglobulin Isotypes Exist, Each with Different Functions
Immunoglobulins can be divided into different classes, or isotypes, based on their distinct biochemical properties. There are two light-chain isotypes, κ and λ. The heavy chains show more variation: in mammals, the major heavy-chain isotypes are µ, δ, γ, α, and ε. These heavy chains can associate with either κ or λ light chains. Depending on the vertebrate species, further subdivisions occur within the α and γ isotypes, and fish possess an isotype that is not found in mammals. The fully assembled immunoglobulin (Ig) derives its name from the heavy chain: µ chains yield IgM; α chains, IgA; γ chains, IgG; δ chains, IgD; and ε chains, IgE. The general structures of the major Ig isotypes are depicted in Figure 23-10. By means of the unique structural features of the Fc portions of their heavy chains, each of the different Ig isotypes carries out specialized functions.
FIGURE 23-10Immunoglobulin isotypes. The different classes of immunoglobulins, called isotypes, may be distinguished biochemically and by immunological techniques. In mice and humans, there are two light-chain isotypes (κ and λ) and five heavy-chain isotypes (µ, δ, γ, ε, α). Each isotype defines a class of immunoglobulin based on the identity of the heavy chain. IgG, IgE, and IgD (not shown) are monomers with generally similar overall structures. IgM and IgA can occur in serum as pentamers and dimers, respectively, accompanied by an accessory subunit, the J chain, in covalent disulfide linkage. This volume-rendered depiction of the immunoglobulins highlights their modular design, with each barrel representing an individual Ig domain. Different isotypes have different functions. See Figure 23-13 for definitions of abbreviations.
The IgM molecule is secreted as a pentamer of H2L2 chains, stabilized by disulfide bonds between the ends of the heavy chains and an additional chain, the J chain. In its pentameric form, IgM possesses ten identical antigen-binding sites (two for each H2L2), which allow high-avidity interactions with surfaces that display the cognate antigen. Avidity is defined as the product of the strength of interactions (affinity) of the available individual binding sites and the number of such binding sites. Many low-affinity interactions can lead to a high-avidity interaction (as in Velcro). Upon its deposition on a surface that carries the cognate antigen, the pentameric IgM molecule assumes a conformation that is highly conducive to activation of the complement cascade, and is thus an effective means of damaging the membrane onto which it is adsorbed and onto which complement proteins are deposited as a consequence.
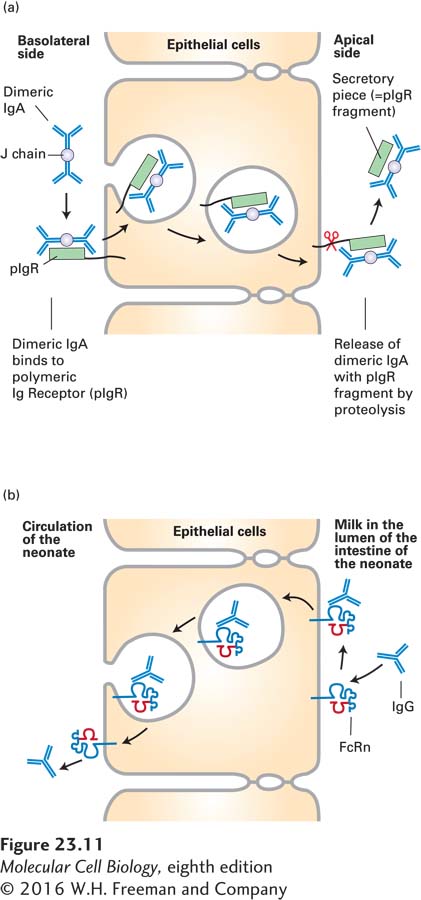
The IgA molecule also interacts with the J chain, forming a dimer of H2L2 molecules. Dimeric IgA can bind to the polymeric IgA receptor found on the basolateral side of epithelial cells, where its binding triggers receptor-mediated endocytosis. Subsequently, the IgA receptor is cleaved, and the dimeric IgA, with a fragment of the receptor (the secretory piece) still attached, is released from the apical side of the epithelial cell. This process, called transcytosis, is an effective means of delivering immunoglobulins from the basolateral side of an epithelium to the apical side (Figure 23-11a). Tears and other secretions, especially in the gastrointestinal tract, are rich in IgA—grams of immunoglobulin are secreted each day!—and so provide protection against environmental pathogens.
FIGURE 23-11Transcytosis of IgA and IgG. (a) IgA, found in tears and in the secretions of various mucous membranes, must be transported across the epithelium. IgA binds to the polymeric IgA receptor and is endocytosed. As the resulting complex is transported across the epithelial monolayer, a portion of the receptor is cleaved, and the IgA, still bound to a portion of the receptor, the secretory piece, is released at the apical side. (b) Suckling rodents acquire Ig from their mother’s milk. At the apical surface of its intestinal epithelium, the newborn possesses the neonatal Fc receptor (FcRn), whose structure resembles that of class I MHC molecules (see Figure 23-23). After this receptor binds to the Fc portion of IgG, transcytosis moves the acquired IgG to the basolateral side of the epithelium. In humans, the syncytial trophoblast in the placenta expresses FcRn and so mediates acquisition of IgG from the maternal circulation and its delivery to the fetus (transplacental transport).
The IgG isotype is important for neutralization of virus particles. This isotype also helps prepare particulate antigens, such as viruses or larger fragments of bacteria, for acquisition by cells equipped with receptors specific for the Fc portion of the IgG molecule.
Page 1091
The immune system of the newborn mammal is immature, but protective antibodies are transferred from the mother to the newborn via the mother’s milk. The receptor responsible for capturing maternal IgG is the neonatal Fc receptor (FcRn), which is present on intestinal epithelial cells in rodents. By transcytosis, maternal IgG captured on the luminal side of the newborn’s intestinal tract is delivered across the gut epithelium and made available for passive protection of the infant rodent (Figure 23-11b). In humans, FcRn is found on fetal cells that contact the maternal circulation in the placenta. Transcytosis of IgG antibodies from the maternal circulation across the placenta delivers maternal antibodies to the fetus. These maternal antibodies will protect the newborn until its immune system is sufficiently mature to produce antibodies on its own. In adults, FcRn is also expressed on endothelial cells and helps control the turnover of IgG in the circulation as well as the delivery of IgG across the endothelial barrier and into underlying tissue.
As we will see in Section 23.3, the IgM and IgD isotypes are expressed as membrane-bound receptors on newly generated B cells. Here the µ chains have an important role in B-cell development and activation.