Each Naive B Cell Produces a Unique Immunoglobulin
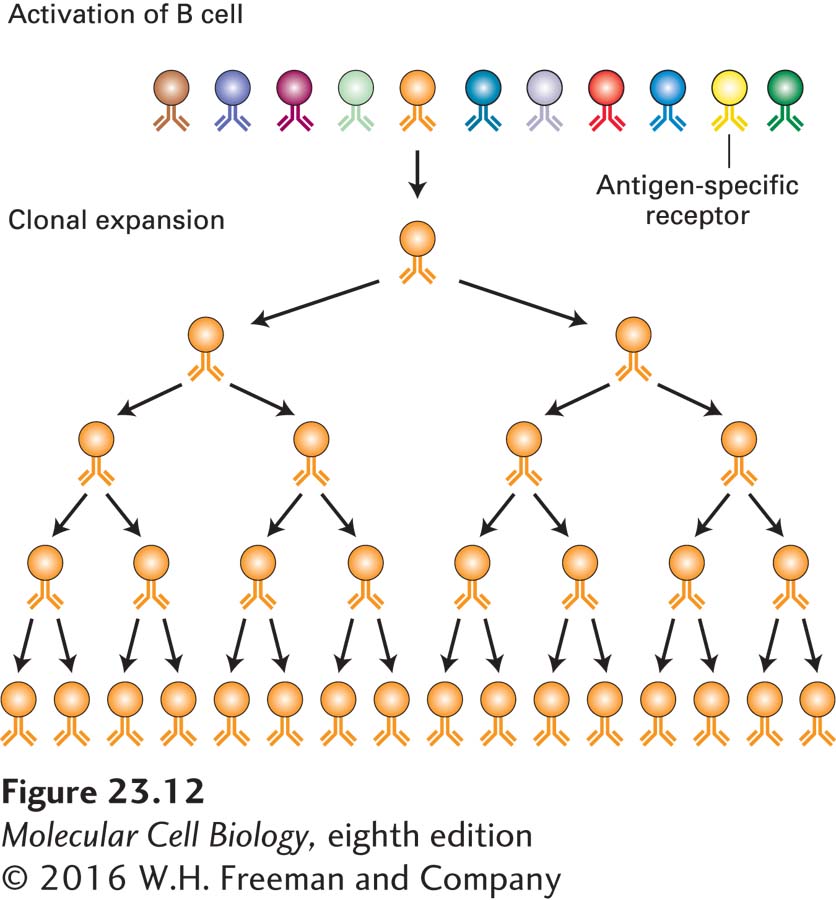
The clonal selection theory stipulates that each naive lymphocyte (not yet having seen its specific antigen) carries an antigen-binding receptor of unique specificity. When a lymphocyte encounters the antigen for which it is specific, clonal expansion (rapid cell division to form a group of cells—a clone—all of which originated from a single precursor cell) occurs and so allows an amplification of the response, culminating in secretion of large amounts of specific antibody (the same one made by the original precursor cell) (Figure 23-12). The antigen-specific antibody is responsible for binding to the antigen and subsequently mediating the clearing of the antigen out of the body. In a typical immune response, the antigen that elicits the response is of complex composition: even the simplest virus contains several distinct proteins, and each protein may present to the immune system several molecularly distinct features that can be recognized independently of one another. Thus many individual lymphocytes respond to a given antigen and expand into independent clones in response to it, each producing its own antigen-specific receptor and antibody of unique structure and therefore with unique binding characteristics (affinity). Because each lymphocyte is endowed with a unique receptor and clonally expands in response to antigen, this response of multiple, independent precursors is characterized as polyclonal.
FIGURE 23-12Clonal selection. The clonal selection theory proposes the existence of a large set of lymphocytes, each equipped with its own unique antigen-specific receptor (indicated by different colors). The antigen that fits with the receptor carried by a particular lymphocyte binds to it and stimulates that lymphocyte to expand clonally. From a modest number of antigen-specific cells, a large number of cells of the desired specificity (and large amounts of their secreted products) may be generated.
B-cell tumors, which represent malignant clonal expansions of individual lymphocytes, enabled the first molecular analysis of the processes that underlie the generation of antibody diversity. A key observation was that tumors derived from lymphocytes may produce large quantities of secreted immunoglobulins. Some of the light chains of these immunoglobulins are secreted in the urine of patients with such tumors. These light chains, called Bence-Jones proteins after their discoverers, can be readily purified and afforded the first target for a protein chemical analysis of immunoglobulins.
Page 1092
Two key observations emerged from this work. First, no two independent tumors produced light chains with identical biochemical properties, suggesting that they were all unique in sequence. Second, the differences in amino acid sequence that distinguished one light chain from another were not randomly distributed, but were clustered in a domain referred to as the variable region of the light chain, or VL. This domain comprises the N-terminal ~110 amino acids of the light chain. The remainder of the sequence is identical for the different light chains (provided they derive from the same isotype, either κ or λ) and is therefore referred to as the constant region, or CL. Immunoglobulins unique to each individual patient were subsequently purified from the patients’ serum. Sequencing of the heavy chains from these preparations revealed that the variable residues that distinguished one heavy chain from another were again concentrated in a well-demarcated domain, referred to as the variable region of the heavy chain, or VH.
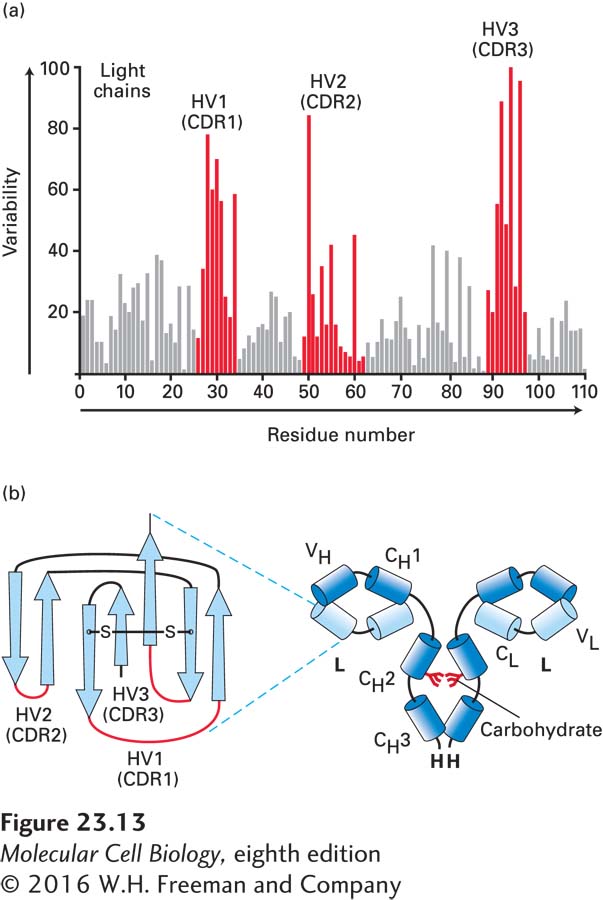
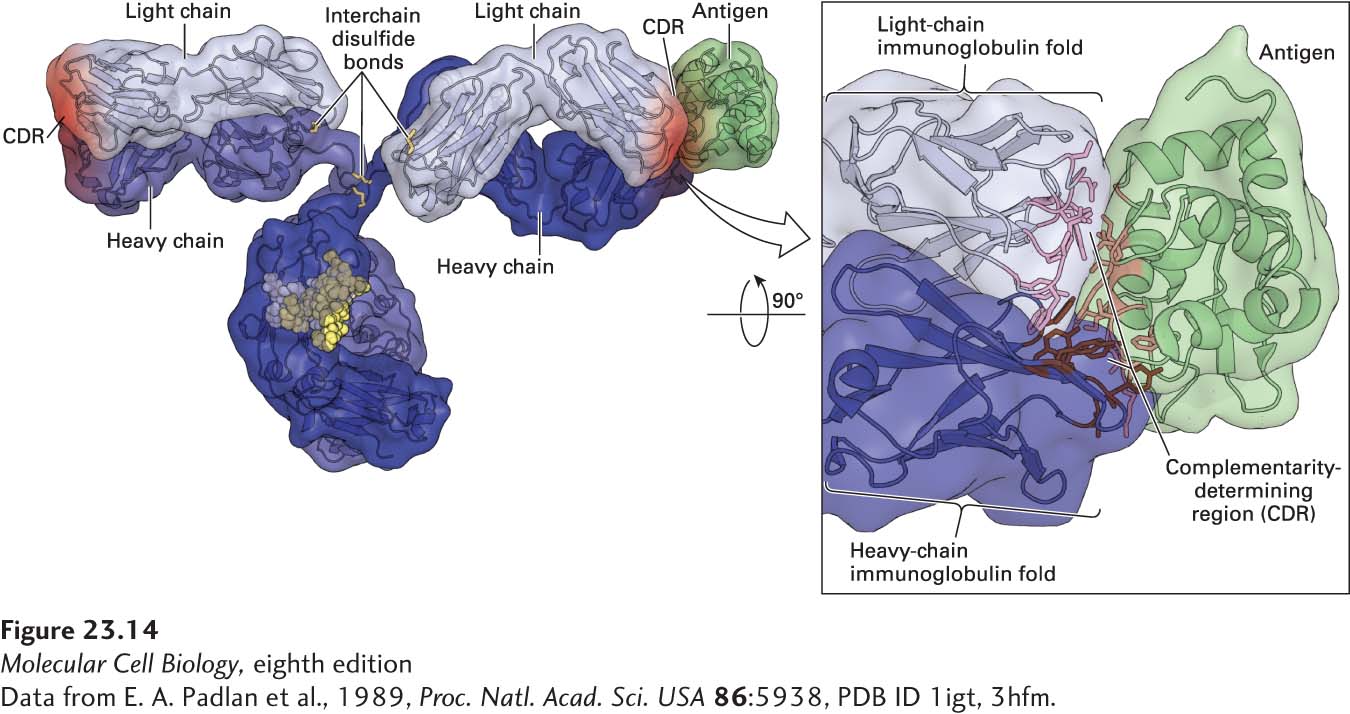
An alignment of variable-region sequences obtained from different light chains showed a nonrandom pattern of regions of variability, revealing three hypervariable regions—HV1, HV2, and HV3—which are sandwiched between what are called framework regions (Figure 23-13a). (Similar alignments for the immunoglobulin heavy-chain sequences also yielded hypervariable regions.) In the properly folded three-dimensional structure of immunoglobulins, these hypervariable regions are in close proximity (Figures 23-13b and 23-14) and make contact with antigen. Thus that portion of an Ig molecule containing the hypervariable regions constitutes the antigen-binding site. For this reason, hypervariable regions are also referred to as complementarity-determining regions (CDRs).
FIGURE 23-13Hypervariable regions and the immunoglobulin fold. (a) Amino acid variability varies with residue position in Ig light chains. Here the percentage of variable-region sequences with variant amino acids is plotted for each position in the sequence. Positions for which many different amino acid side chains are present are assigned a high variability index; those that are invariant among the sequences compared are assigned a value of 0. This analysis reveals three regions of increased variability: hypervariability (HV) regions 1, 2, and 3; these regions are also called complementarity-determining regions (CDRs). (b) Volume-rendered depiction of F(ab′)2 fragment (right) and ribbon diagram of a typical Ig light-chain variable region (VL) with the positions of the hypervariable regions indicated in red (left). The hypervariable regions are found in the loops that connect the β strands and make contact with antigen. The β strands (rendered as arrows) make up two β sheets and constitute the framework region. Each variable and constant domain has this characteristic three-dimensional structure, called the immunoglobulin fold. L = light chain; H = heavy chain; VH = heavy-chain variable region; VL = light-chain variable region; CH1, CH2, CH3 = heavy-chain constant domains; CL = light-chain constant region.
[Data from E. A. Padlan et al., 1989, Proc. Natl. Acad. Sci. USA86:5938, PDB ID 1igt, 3hfm.]
FIGURE 23-14Immunoglobulin structure. This model shows the three-dimensional structure of an immunoglobulin molecule complexed with hen egg-white lysozyme (a protein antigen) as determined by x-ray crystallography.
[Data from E. A. Padlan et al., 1989, Proc. Natl. Acad. Sci. USA86:5938, PDB ID 1igt, 3hfm.]
Page 1093
The difficulty of encoding directly in the inherited genome (germ line) all the information necessary to generate the enormously diverse antibody repertoire (more than a million different antibody molecules in what we now know is a genome encoding about 20,000 independent genes) led to suggestions of unique genetic mechanisms to account for this diversity. Given the size of a typical antibody heavy chain and light chain (each heavy chain–light chain combination, if encoded as such, would require 2.5–3.5 kb of DNA, depending on the isotype), it is immediately obvious that encoding a set of antibody molecules of sufficient diversity to provide adequate protection against the wide array of pathogens and other foreign substances to which an organism is exposed would rapidly exhaust its DNA coding capacity. We shall see that, indeed, unique mechanisms are at work to create an adequately diverse set of antibodies.