The Class I MHC Pathway Presents Cytosolic Antigens
Figure 23-25 summarizes the six steps in the class I MHC pathway using a virus-infected cell as an example.
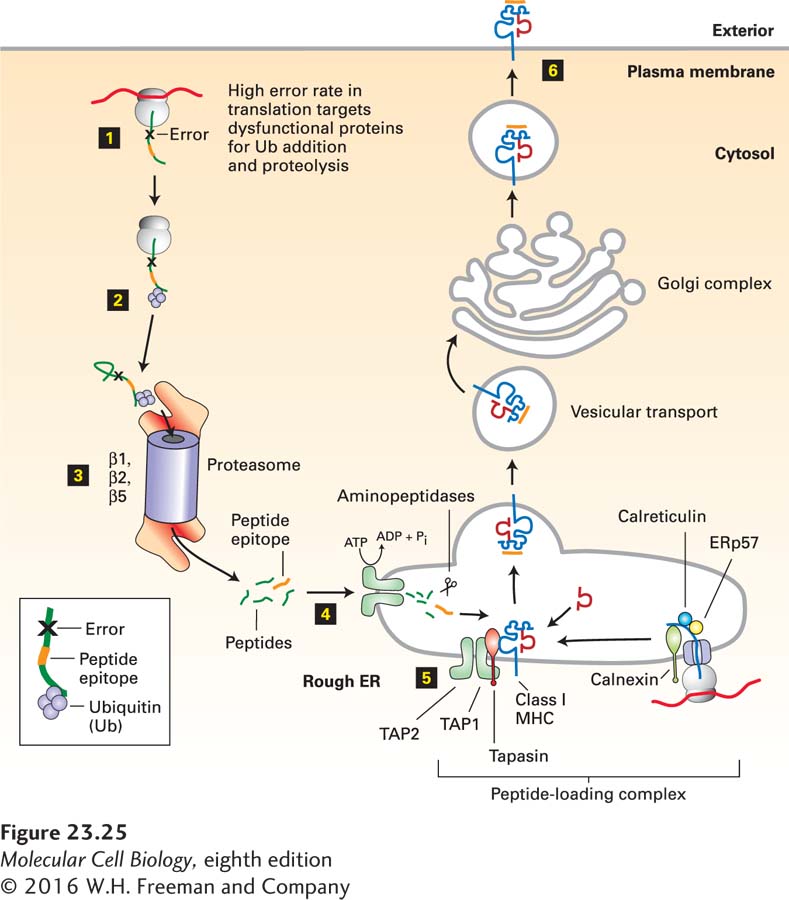
FIGURE 23-25Class I MHC pathway of antigen processing and presentation. Step 1: Acquisition of antigen is synonymous with the production of proteins with errors (premature termination, misincorporation). Step 2: Dysfunctional proteins are targeted for degradation by ubiquitinylation. Step 3: Proteolysis is carried out by the proteasome. In cells exposed to interferon γ, the catalytically active β subunits of the proteasome are replaced by interferon-induced immune-specific β subunits. Step 4: Peptides are delivered to the interior of the endoplasmic reticulum (ER) via the dimeric TAP peptide transporter. Step 5: Peptide is loaded onto newly made class I MHC molecules within the peptide-loading complex. Step 6: The fully assembled class I MHC–peptide complex is transported to the cell surface via the secretory pathway. See text for details.
1Acquisition of Antigen: In the case of a viral infection, acquisition of antigen is usually synonymous with the infected state. Viruses rely on host biosynthetic pathways to generate new viral proteins. Protein synthesis, unlike DNA replication, is an error-prone process, in which a fraction of newly initiated polypeptide chains are terminated prematurely or suffer from other errors (misincorporation of amino acids, frameshifts, improper or delayed folding). These mistakes in protein synthesis affect the host cell’s own proteins and those specified by viral genomes equally. Such error-containing proteins must be rapidly removed so as not to clog up the cytoplasm, engage partner proteins in nonproductive interactions, or even act as dominant negative versions of a protein. Properly folded proteins may also sustain damage that leads to their unfolding, completely or in part, and necessitates their removal. The rate of cytosolic proteolysis of these dysfunctional proteins must be matched to the rate at which mistakes in protein synthesis and folding occur. These proteins are an important source of the peptides destined for presentation by class I MHC molecules. With the exception of a specialized process called cross-presentation (discussed below), the class I MHC pathway results in the formation of peptide-MHC complexes in which the peptides are derived from proteins synthesized by the class I MHC-bearing cell itself.
2Targeting Antigen for Destruction: For the most part, polyubiquitinylation is responsible for targeting a protein for destruction (see Chapter 3, page 99). Polyubiquitinylation is a covalent modification that is tightly regulated.
Page 1111
3Proteolysis: Polyubiquitinylated proteins are destroyed by proteolysis in proteasomes. The proteasome is a protease that engages its substrates and, without the release of intermediates, yields peptides in the size range of 3–20 amino acids as its final digestion products (see Figure 3-31). During the course of an inflammatory response and in response to interferon γ, the three catalytically active β subunits (β1, β2, β5) of the proteasome can be replaced by three immune-specific subunits: β1i, β2i, and β5i. The β1i, β2i, and β5i subunits are encoded in the MHC region of the genome. The net result of this replacement is the generation of an immunoproteasome, the output (length of peptide products) of which is matched to the requirements for peptide binding by class I MHC molecules. The immunoproteasome adjusts the average length of the peptides produced as well as the sites at which cleavage occurs. Given the central role of the proteasome in the generation of the peptides presented by class I MHC molecules, proteasome inhibitors interfere potently with antigen processing via the class I MHC pathway.
4Delivery of Peptides to Class I MHC Molecules: Protein synthesis, polyubiquitinylation, and proteasomal proteolysis all occur in the cytoplasm, whereas peptide binding by class I MHC molecules occurs in the lumen of the endoplasmic reticulum (ER). Thus peptides must cross the ER membrane to gain access to class I molecules, a process mediated by the heterodimeric TAP complex, a member of the ABC superfamily of ATP-powered pumps (see Figure 11-15). The TAP complex binds peptides on the cytoplasmic face of the ER and, in a cycle that includes ATP binding and hydrolysis, translocates them into the ER. The specificity of the TAP complex is such that it can transport only a subset of all cytosolic peptides, primarily those in the length range of 5–10 amino acids, that are compatible with the circumscribed length of peptides that can fit into the class I MHC molecules. The mouse TAP complex shows a pronounced preference for peptides that terminate in leucine, valine, isoleucine, or methionine residues, which match the binding preference of class I MHC molecules. The genes encoding the TAP1 and TAP2 subunits composing the TAP complex are located in the MHC region.
5Binding of Peptides to Class I MHC Molecules: Within the ER, newly synthesized class I MHC molecules are part of a multiprotein complex referred to as the peptide-loading complex. This complex includes two chaperones (calnexin and calreticulin) and the oxidoreductase Erp57. Another chaperone (tapasin) interacts with both the TAP complex and the class I MHC molecule about to receive peptide. The physical proximity of TAP and the class I MHC molecule is maintained by tapasin. Once peptide loading onto the class I MHC molecule has occurred, a conformational change releases the loaded class I MHC molecule from the peptide-loading complex. This arrangement effectively ensures that only peptide-loaded class I MHC molecules are released from the ER and then transported to and displayed at the cell surface. The overall efficiency of this pathway is such that approximately 4000 molecules of a given protein must be destroyed to generate a single MHC-peptide complex carrying a peptide from that particular polypeptide.
6Display of Class I MHC–Peptide Complexes at the Cell Surface: Once peptide loading is complete, the class I MHC–peptide complex is released from the peptide-loading complex and enters the constitutive secretory pathway (see Figure 14-1). Transfer from the Golgi to the cell surface is rapid and completes the biosynthetic pathway of a class I MHC–peptide complex.
The entire sequence of events in the class I pathway occurs constitutively in all nucleated cells, all of which express class I MHC molecules and the other required proteins, or can be induced to do so. As we have seen, exposure to cytokines such as interferon γ can induce immune-specific proteasomal subunits to generate immunoproteasomes with enhanced ability to produce the appropriate peptides for presentation by class I MHC molecules. In the absence of a viral infection, protein synthesis and proteolysis continuously generate a stream of peptides that are loaded onto class I MHC molecules. Healthy, normal cells therefore display on their surfaces a representative selection of peptides derived from their own proteins. There may be several thousand distinct MHC-peptide combinations displayed at the surface of a typical nucleated cell. The display of MHC–self-peptide complexes on the surfaces of normal, uninfected cells plays an essential role in the immune system. It is not until a virus makes its appearance that virus-derived peptides begin to make a contribution to the display of peptide-MHC complexes on cell surfaces.
As we noted above, a properly functioning immune system must be able to distinguish self (nonpathogenic) antigens from nonself (potentially pathogenic) antigens. The small organ called the thymus—located near the sternum at the level of the heart in humans—plays a critical role in controlling the ability of the immune system to identify self and nonself. Developing T cells in the thymus, referred to as thymocytes, calibrate their antigen-specific receptors to the sets of MHC-peptide complexes generated on thymic epithelial cells and learn to recognize self-MHC products as the guideposts—or restriction elements, in immunological parlance—on which they must henceforth rely for antigen recognition. At the same time, the display of self peptides by self MHC molecules in the thymus enables the developing T cell to learn which peptide-MHC combinations are self-derived and must therefore be ignored to avoid a self-destructive autoimmune reaction. T-cell development is thus driven by self MHC molecules loaded with self peptides, a “template” on which a useful repertoire of T cells can be molded. Simply put, any T cell that bears a receptor that too strongly reacts with self-MHC–self-peptide complexes is potentially dangerous when it leaves the thymus and must be eliminated. This process of selection will be discussed below.
An exception to the usual mode of antigen presentation that is nonetheless crucial in the development of cytotoxic T cells is cross-presentation. This term refers to the acquisition by dendritic cells of apoptotic cell remnants, complexes composed of antigen bound to antibody, and possibly other forms of antigen, by phagocytosis. By a pathway that has yet to be fully understood, these materials escape from phagosomal or endosomal compartments into the cytosol, where they are then handled according to the steps described above. Dendritic cells are the most efficient at cross-presentation, and so allow the loading of class I MHC molecules complexed with peptides that derive from cells other than the APC itself.