The Class II MHC Pathway Presents Antigens Delivered to the Endocytic Pathway
Although class I MHC and class II MHC molecules show a striking structural resemblance, the manner in which the two classes acquire peptide and their function in antigen recognition differ greatly. Whereas the primary function of class I MHC molecules is to guide CD8-bearing cytotoxic T cells to their target (usually infected) cells, class II MHC molecules serve to guide CD4-bearing helper T cells to the cells with which they interact, primarily professional APCs. Activated helper T cells provide protection not only by helping B cells to produce antibodies, but also by means of the complex sets of cytokines they produce, which activate phagocytic cells to clear pathogens or help set up an inflammatory response.
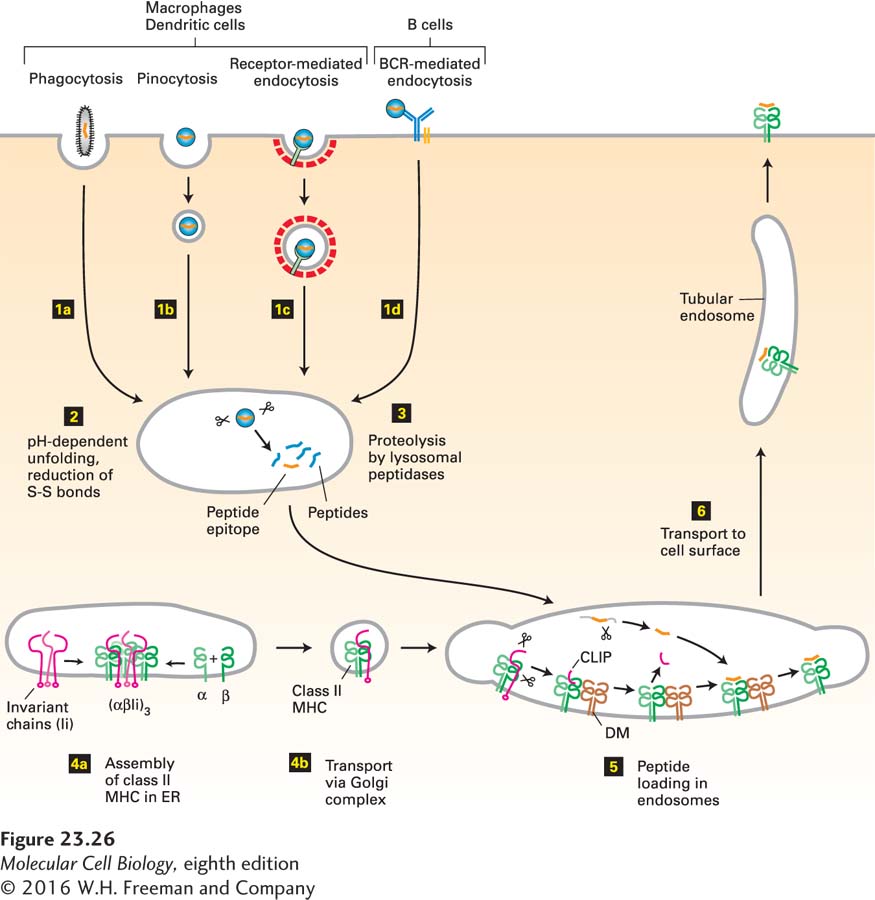
As noted previously, class II MHC molecules are expressed primarily by professional APCs: dendritic cells and macrophages, which are phagocytic, and B cells, which are not. Hence the class II MHC pathway of antigen processing and presentation generally occurs only in these cells. The steps in this pathway are depicted in Figure 23-26.
FIGURE 23-26Class II MHC pathway of antigen processing and presentation. Step 1: Particulate antigens are acquired by phagocytosis and nonparticulate antigens by pinocytosis or endocytosis. Step 2:Exposure of antigen to the acidic and reducing environment of endosomes and lysosomes prepares the antigen for proteolysis. Step 3:The antigen is broken down by various proteases in endosomal and lysosomal compartments. Step 4: Class II MHC molecules, assembled in the ER from their subunits, are delivered to endosomal and lysosomal compartments by means of signals contained in the associated invariant (Ii) chain. This delivery targets late endosomes, lysosomes, and early endosomes, ensuring that class II MHC molecules are exposed to the products of proteolytic breakdown of antigen along the entire endocytic pathway. Step 5: Peptide loading is accomplished with the assistance of DM, a class II MHC–like chaperone protein. Step 6: Peptide-loaded class II MHC molecules are displayed at the cell surface. See text for details.
Page 1113
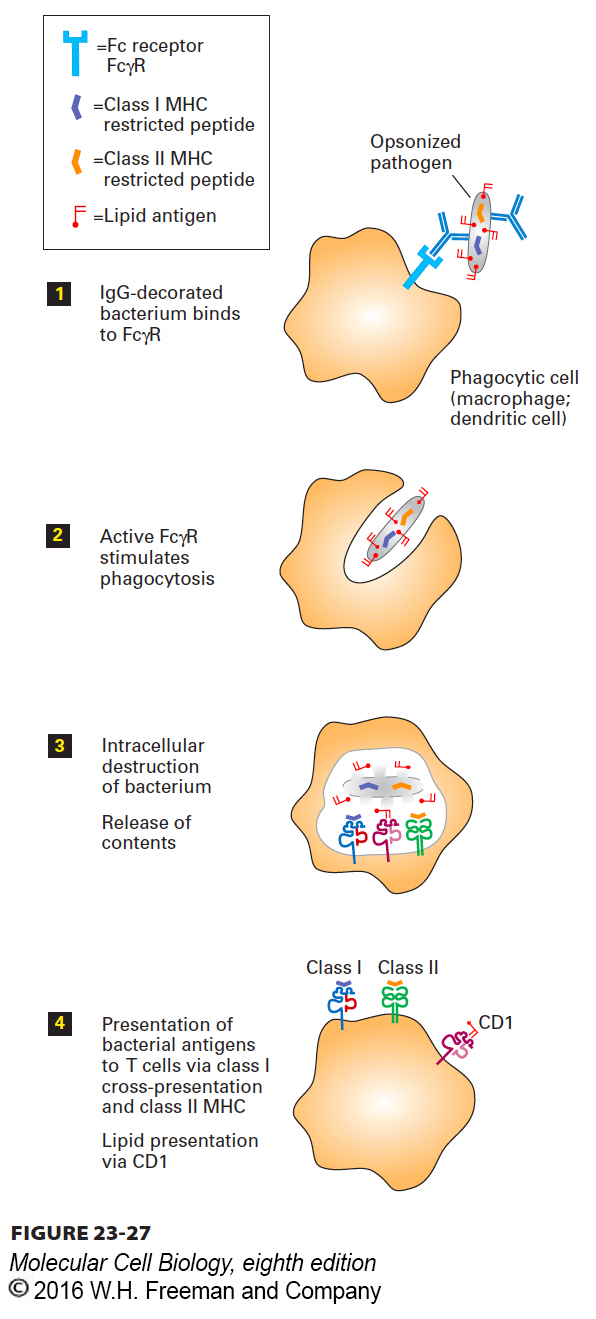
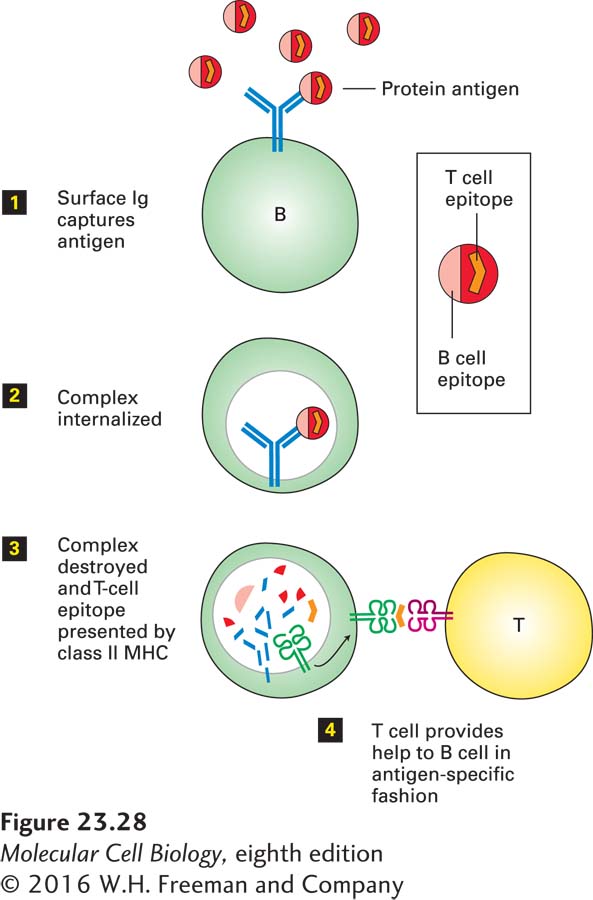
1Acquisition of Antigen: In the class II MHC pathway, antigen is acquired by pinocytosis, phagocytosis, or receptor-mediated endocytosis. Pinocytosis, which is rather nonspecific, involves the delivery, by a process of membrane invagination and fission, of a volume of extracellular fluid and the molecules dissolved therein. Phagocytosis, the ingestion of particulate materials such as bacteria, viruses, and remnants of dead cells, involves extensive remodeling of the actin-based cytoskeleton to accommodate the incoming particle. Although phagocytosis may be initiated by specific receptor-ligand interactions, these are not always required: even latex particles and other particulates such as glass beads can be ingested very efficiently by macrophages. Pathogens decorated by antibodies and certain complement components are targeted to macrophages and dendritic cells, which recognize them by means of cell-surface receptors for complement components or for the Fc portion of immunoglobulins, then phagocytose them (Figure 23-27). Macrophages and dendritic cells also express several types of less selective receptors (e.g., C-type lectins, Toll-like receptors, scavenger receptors) that recognize molecular patterns in both soluble and particulate antigens; these cells then internalize the bound antigens by receptor-mediated endocytosis. B cells, which are not phagocytic, can also acquire antigens by receptor-mediated endocytosis using their antigen-specific B-cell receptors (Figure 23-28). Finally, cytosolic antigens may enter the class II MHC pathway via autophagy (see Figure 14-35).
FIGURE 23-27Presentation of opsonized antigen by phagocytic cells. By means of Fc receptors such as FcγR displayed on their cell surface, specialized phagocytic cells such as macrophages or dendritic cells can bind and ingest pathogens that have been decorated with antibodies (opsonization). After digestion of the phagocytosed particle (e.g., immune complex, bacterium, virus), some of the peptides produced, including fragments of the pathogen (orange), are loaded onto class II MHC molecules (green). Class II MHC–peptide complexes displayed at the surface allow activation of T cells specific for these MHC-peptide combinations. Lipid antigens are delivered to the class I MHC–like molecule CD1 (pink), whose binding site is specialized to accommodate lipids. Certain pathogen-derived peptides (purple) may be delivered to class I MHC products (blue) by means of cross-presentation. The mechanisms that underlie cross-presentation remain to be clarified.
FIGURE 23-28Antigen presentation by B cells. B cells bind antigen, even if present at low concentration, to their B-cell receptors, or surface Ig. The immune complex that results is internalized and then delivered to endosomal or lysosomal compartments, where it is destroyed. Peptides liberated from the immune complex, including fragments of the protein antigen, are displayed as class II MHC–peptide complexes at the cell surface. Helper T cells specific for the displayed complex can now provide help to the B cell. This help is MHC restricted and antigen specific.
2Targeting Antigen for Destruction: Proteolysis is required to convert intact protein antigens into peptides of a size suitable for binding to class II MHC molecules. Protein antigens are targeted for degradation by progressive unfolding, brought about by the drop in pH as proteins progress along the endocytic pathway. The pH of the extracellular environment is around pH 7.2, and that in early endosomes is between pH 6.5 and 5.5; in late endosomes and lysosomes the pH may drop to pH 4.5. ATP-powered V-class proton pumps in the endosomal and lysosomal membranes are responsible for this acidification (see Figure 11-9). Proteins that are stable at neutral pH tend to unfold when they are exposed to extremes of pH through rupture of hydrogen bonds and destabilization of salt bridges. Furthermore, the environment in the endosomal or lysosomal compartment is a reducing one, in which lysosomes attain a concentration of reducing equivalents in the millimolar range. Reduction of the disulfide bonds that stabilize many extracellular proteins can also be catalyzed by a thioreductase inducible by exposure to interferon γ. The combined action of low pH and reducing environment prepares the antigens for proteolysis.
3Proteolysis: Degradation of proteins in the class II MHC pathway is carried out by a large set of lysosomal proteases, collectively referred to as cathepsins, which are either cysteine or aspartyl proteases. Other proteases, such as asparagine-specific endoprotease, may also contribute to proteolysis. A wide range of peptide fragments is produced, including some that can bind to class II MHC molecules. The lysosomal proteases operate optimally at the acidic pH within lysosomes. Consequently, agents that inhibit the activity of the V-class proton pumps that maintain their acidification interfere with antigen processing, as do inhibitors of lysosomal proteases.
Page 1114
4Encounter of Peptides with Class II MHC Molecules: Recall that most class II MHC molecules synthesized in the endoplasmic reticulum are directed to late endosomes. The peptides generated by proteolysis reside in the same topological space as the class II MHC molecules themselves—they do not have to cross a membrane, as is the case for peptides destined to bind to class I MHC molecules (see Figure 23-25). To allow peptides and class II MHC molecules to meet, the (αβIi)3 complex is transported via the secretory pathway to endosomal compartments.
5Binding of Peptides to Class II Molecules: The (αβIi)3 complex delivered to endosomal compartments is incapable of binding peptide because the peptide-binding cleft in the class II molecule is occupied by the invariant chain (Ii). For the same reason, newly assembled (αβIi)3 complexes do not compete for class I MHC–destined peptides delivered to the ER via TAP: their peptide-binding site is already occupied by Ii. Recall that the ER is where both class I and class II MHC molecules assemble. The presence of Ii in the nascent class II MHC complex ensures that class II MHC molecules do not bind peptide in the ER. The same proteases in endosomes and lysosomes that act on internalized antigens and degrade them into peptides also act on the (αβIi)3 complex, resulting in removal of the Ii molecule from the complex with the exception of a small portion called the CLIP segment. Because it is firmly lodged in the class II MHC peptide-binding cleft, CLIP is resistant to proteolytic attack. The class II MHC molecules themselves are also resistant to unfolding and proteolytic attack under the conditions that prevail in the endocytic pathway. The CLIP segment is removed from the αβ heterodimer by the chaperone DM. The newly vacated peptide-binding cleft of the class II MHC molecule may now bind the peptides that are abundantly present in the endocytic pathway. Although the DM protein is MHC encoded and structurally very similar to class II molecules, it does not itself bind peptides. However, newly formed class II MHC–peptide complexes are themselves susceptible to further “editing” by DM, which may dislodge the peptide already bound, until the class II molecule acquires a peptide that binds so strongly that it cannot be removed by DM. The resulting class II MHC–peptide complexes are extremely stable, with estimated half-lives in excess of 24 hours.
6Display of Class II MHC–Peptide Complexes at the Cell Surface: The newly generated class II MHC–peptide complexes are localized mostly in late endosomal compartments, which include multivesicular endosomes (or bodies) (see Figure 14-33). Recruitment of the internal vesicles of the multivesicular bodies to the delimiting membrane expands their surface area: by formation of tubular membranes, laid down along tracks of microtubules, these compartments elongate and ultimately deliver class II MHC–peptide complexes to the surface by membrane fusion. These events are tightly regulated: tubulation and delivery of class II MHC molecules to the surface are enhanced in dendritic cells and macrophages following their activation by signals generated in response to infection, such as bacterial lipopolysaccharide, which is detected by Toll-like receptors on the surfaces of these professional APCs, as well as inflammatory cytokines, such as interferon γ, produced by CD4-expressing helper T cells.
For professional APCs, the above steps are constitutive—happening all the time—but they can be modulated by exposure to microbial agents and cytokines. In addition to the pathways described here for class I and class II MHC products, there is a category of class I MHC–related molecules, the CD1 proteins, that are specialized in the presentation of lipid antigens. The structure of a CD1 molecule resembles that of a class I MHC molecule: a larger subunit complexed with β2-microglobulin. Many species of bacteria produce lipids whose chemical structures are not found in their mammalian hosts. These lipids can serve as antigens when presented by CD1 molecules, to which they bind via a lipid-binding pocket that is conceptually similar to that of most MHC molecules. Signals in the cytoplasmic tail of the larger CD1 subunit target these molecules to endosomal or lysosomal compartments, where loading with antigenic lipids occurs. The CD1-lipid complexes engage a relatively rare class of T cells, referred to as NKT cells, as well as γδ T cells, both described below. NKT cells fulfill an important role in cytokine production and help initiate and orchestrate adaptive immune responses via their cytokine outputs.