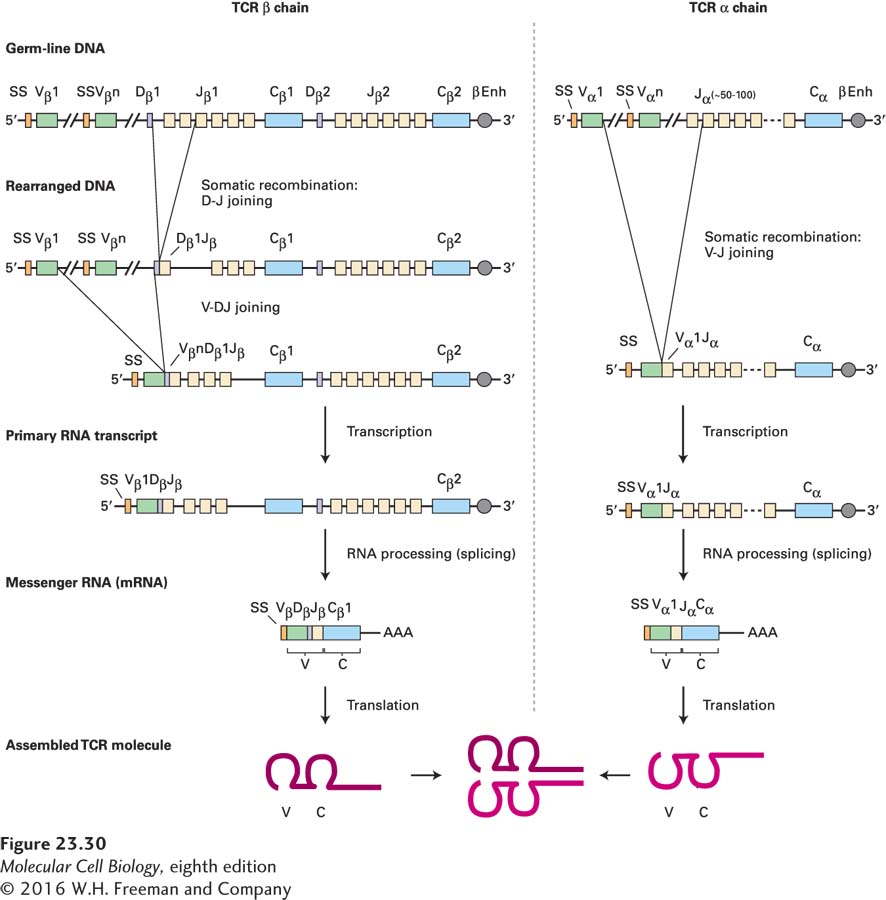
TCR Genes Are Rearranged in a Manner Similar to Immunoglobulin Genes
Virtually all antigen-specific receptors generated by somatic recombination contain a subunit that is the product of V-D-J recombination (e.g., Ig heavy chain; TCR β chain) and another that is the product of V-J recombination (e.g., Ig light chain; TCR α chain). The mechanism of V-D-J and V-J recombination for TCRs is essentially identical to that described for immunoglobulin genes and requires all the component proteins composing the nonhomologous end-joining machinery: RAG1, RAG2, Ku70, Ku80, the catalytic subunit of DNA-dependent protein kinase, XRCC4, DNA ligase IV, and Artemis. Recombination signal sequences (RSSs) are required, and recombination obeys the 12/23-bp spacer rule (Figure 23-30).
FIGURE 23-30Organization and recombination of TCR loci. The organization of TCR loci is in principle similar to that of immunoglobulin loci (see Figure 23-15). Left: The TCR β-chain locus includes a cluster of V segments, a cluster of D segments, and several J segments, downstream of which are two constant regions. The arrangement of the recombination signals is such that not only is D-J joining allowed, but also V-D-J joining. Direct V-J joining in the TCRβ locus is not observed. Right: The TCR α-chain locus is composed of a cluster of V segments and a large number of J segments. SS = exon encoding signal sequence; Enh = enhancer.
A number of noteworthy features characterize the organization and rearrangement of the TCR loci. First, the organization of the RSSs is such that D-to-D rearrangements are allowed, unlike the case for Ig. Second, terminal deoxynucleotidyltransferase (TdT) is active at the time the TCR genes are rearranged, and therefore N nucleotides can be present in all rearranged TCR genes. Third, in humans and mice, the TCR δ locus is embedded within the TCR α locus. This organization results in complete excision of the interposed δ locus when TCRα rearrangement occurs, so a choice of the TCR α locus for rearrangement precludes use of the δ locus, which is lost by deletion. T cells that express the αβ receptor and those that express the γδ receptor are considered separate lineages with distinct functions. Among the T cells expressing γδ receptors are some capable of recognizing the CD1 molecule, which is specialized for the presentation of lipid antigens. The γδ T cells are programmed to home in on distinct anatomic sites (e.g., the epithelium lining the genital tract, the skin) and probably play a role in host defense against pathogens commonly found at these sites.
Page 1117
Deficiencies in the key components of the recombination apparatus, such as the RAG recombinases, preclude rearrangement of TCR genes. As we have seen for B cells, development of lymphocytes is strictly dependent on the rearrangement of the antigen-receptor genes. A deficiency in either RAG1 or RAG2 thus prevents both B-cell and T-cell development. Mice with homozygous RAG gene knockouts are frequently used to assess the roles of B and T cells in physiological and pathophysiological processes.