Production of High-Affinity Antibodies Requires Collaboration Between B and T cells
To generate the high-affinity antibodies that are necessary for tight binding to antigens and effective neutralization of pathogens, B cells require assistance from T cells. B-cell activation requires both a source of antigen to engage the BCR and the presence of activated antigen-specific T cells.
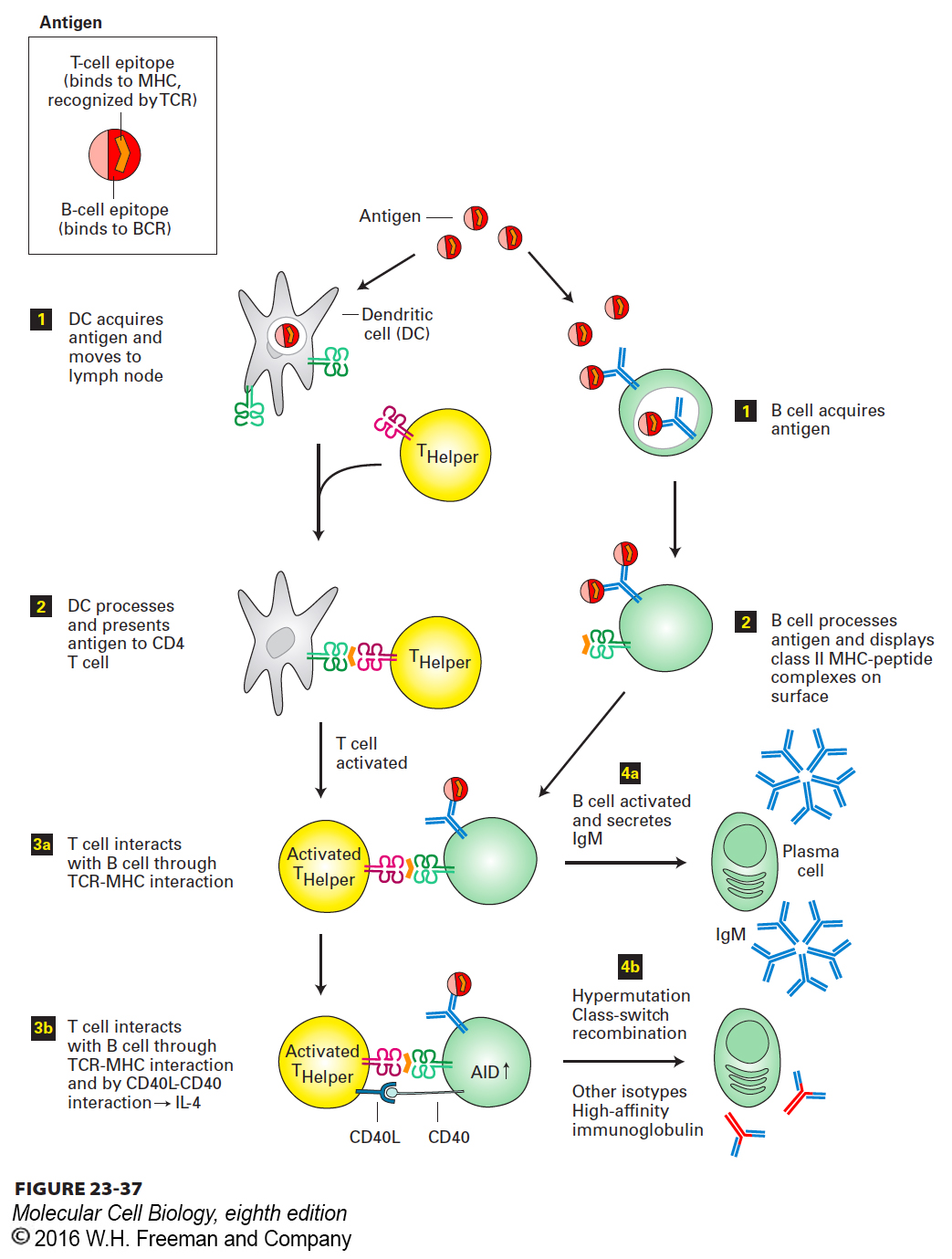
Soluble antigen reaches the lymph node from the periphery by transport through the afferent lymphatic vessels (see Figure 23-7). Bacterial growth is accompanied by the release of microbial products that can serve as antigens. If the infection is accompanied by local tissue destruction, activation of the complement cascade results in the killing of bacteria and the concomitant release of bacterial proteins, which are also delivered via the lymphatic vessels to the draining lymph node. Antigens covalently modified by proteins of the complement system are superior to their unmodified counterparts in the activation of B cells through engagement of complement receptors on those cells, which serve as co-receptors for the B-cell receptor. B cells that acquire antigen via their BCRs internalize the antibody-antigen complex by endocytosis and process it for presentation via the class II MHC pathway. B cells that have successfully engaged antigen thus convert it into a call for T-cell help in the form of a class II MHC–peptide complex expressed on the cell surface (Figure 23-37, step 2). Note that the peptide on the antigen molecule recognized by the B-cell receptor may be quite distinct from the peptide ultimately displayed on the cell surface in association with a class II MHC molecule. As long as the B-cell epitope and the class II MHC–presented peptide—a T-cell epitope—are physically linked, successful B-cell differentiation and antibody production can be initiated.
FIGURE 23-37Collaboration between T and B cells is required to initiate the production of antibodies.Left: Activation of T cells by means of antigen-loaded dendritic cells (DCs). Right: Antigen acquisition by and subsequent activation of B cells. Step 1: Professional antigen-presenting cells (dendritic cells, B cells) acquire antigen. Step 2: Professional APCs internalize and process antigen. T-cell activation occurs when dendritic cells present antigen to T cells. Step 3a: Activated T cells engage antigen-experienced B cells through peptide-MHC complexes displayed on the surface of the B cell. Step 3b: T cells that are persistently activated initiate expression of the CD40 ligand (CD40L), a prerequisite for B cells becoming fully activated and turning on the enzymatic machinery (AID) to initiate class-switch recombination and somatic hypermutation. Step 4a: A B cell that receives the appropriate instructions from CD4 helper T cells becomes an IgM-secreting plasma cell. Step 4b: A B cell that receives signals from activated CD4 helper T cells in the form of CD40–CD40L interactions and the appropriate cytokines can switch to other immunoglobulin isotypes and engage in somatic hypermutation.
This concept of linked recognition—namely, the engagement of antigen by the B cell’s BCR and the display of antigen-derived fragments to T cells by class II MHC molecules—explains why there is a minimum size for molecules that can be used to successfully elicit a high-affinity antibody response, as we will see below. Such immunogenic molecules must fulfill several criteria: they must contain the epitope that binds to the B-cell receptor, they must undergo endocytosis and proteolysis, and a proteolyzed fragment of the protein must bind to the allelic class II MHC molecules available in order to be presented as a class II MHC–peptide complex, which serves as a call for T-cell help.
Often investigators would like to generate an antibody (either polyclonal or monoclonal) that can recognize a short peptide fragment from a larger protein. Such antibodies can be used for a variety of experiments, including detection of a target molecule by immunofluorescence or immunoprecipitation. These antibodies are called anti-peptide antibodies. If the peptide alone is used as an immunogen (injected into an animal [e.g., a rabbit, goat, or mouse] to generate antibodies), it probably will not successfully induce robust antibody formation, even though there may be B cells with BCRs that can bind tightly to the peptide. The reason is that it is unlikely that those B cells will be able to generate a complex of a class II MHC with that same peptide that can recruit helper T cells to drive proliferation and affinity maturation. For this reason, synthetic peptides used to elicit antibodies are conjugated to carrier proteins to improve their immunogenicity; the carrier proteins serve as the source of peptides for presentation via class II MHC products. Only through recognition of such a class II MHC–peptide complex via its T-cell receptor can a T cell provide the help necessary for the B cell to run its complete course of differentiation leading to robust, high-affinity antibody production.
This concept applies equally to B cells capable of recognizing particular modifications on proteins or peptides. Antibodies that recognize the phosphorylated form of a kinase are commonly raised by immunization of experimental animals with the phosphorylated peptide in question, conjugated to a carrier protein. An appropriately specific B cell recognizes the phosphorylated site on the peptide of interest, internalizes the phosphorylated peptide and carrier, and generates a complex set of peptides by endosomal proteolysis of the carrier protein. Among these peptides, there should be at least one that can bind to the class II MHC molecules carried by that B cell. If properly displayed at the surface of the B cell, this class II MHC–peptide complex becomes the call for T-cell help, which is provided by helper T cells equipped with receptors capable of recognizing the complex of class II MHC molecule and carrier-derived peptide.
The helper T cell identifies, via its TCR, an antigen-experienced B cell by means of the class II MHC–peptide complex the B cell displays. The B cell also displays co-stimulatory molecules and receptors for cytokines produced by the activated T cell (e.g., IL-4). After interacting with T cells, these B cells proliferate. Some of them differentiate into plasma cells; others are set aside and become memory B cells. The first wave of antibodies they produce is always IgM. Class switching to other isotypes and somatic hypermutation (necessary for the generation and selection of high-affinity antibodies) require the persistence of antigen or repeated exposure to antigen. In addition to cytokines, B cells require cell-cell contacts to initiate these processes. These contacts involve CD40 protein on B cells and CD40L on T cells. These proteins are members of the TNF–TNF receptor family. Recent work on HIV suggests that extensive hypermutation is a prerequisite for the generation of broadly neutralizing antibodies (antibodies that can neutralize a broad selection of highly variable HIV isolates). More insight in the control of somatic hypermutation will be required to understand the nature of the antigen(s) capable of eliciting such desirable antibodies as a prophylactic strategy.