Gain-of-Function Mutations Convert Proto-oncogenes into Oncogenes
Any gene that encodes a protein able to transform cells in culture, usually in combination with other cell alterations, or to induce cancer in animals is considered to be an oncogene. Of the many known oncogenes, all but a few are derived from normal cellular genes (i.e., proto-oncogenes) whose wild-type products promote cell proliferation or other features important to cancer. For example, the RAS gene discussed previously is a proto-oncogene that encodes an intracellular signal-transducing protein that promotes cell division; the mutant rasD gene derived from RAS is an oncogene whose protein product provides an excessive or uncontrolled proliferation-promoting signal. Other proto-oncogenes encode growth-promoting signaling molecules and their receptors, anti-apoptotic (cell-survival) proteins, and transcription factors.
Page 1150
Page 1151
Conversion of a proto-oncogene into an oncogene, also called activation, generally involves a gain-of-function mutation. At least four mechanisms can produce oncogenes from the corresponding proto-oncogenes:
A point mutation (i.e., a change in a single base pair) in a proto-oncogene that results in a hyperactive or constitutively active protein product
A chromosomal translocation that fuses two genes together to produce a hybrid gene encoding a chimeric protein whose activity, unlike that of the parent proteins, is constitutive
A chromosomal translocation that brings a growth regulatory gene under the control of alternative enhancers that cause inappropriate expression of the gene
Amplification (i.e., abnormal DNA replication) of a DNA segment including a proto-oncogene so that numerous copies exist, leading to overproduction of the encoded protein
An oncogene formed by either of the first two mechanisms encodes an oncoprotein that differs from the normal protein encoded by the corresponding proto-oncogene. In contrast, the other two mechanisms generate oncogenes whose protein products are identical to the normal proteins; their oncogenic effect is due to their production at higher than normal levels or in cells where they are not normally produced.
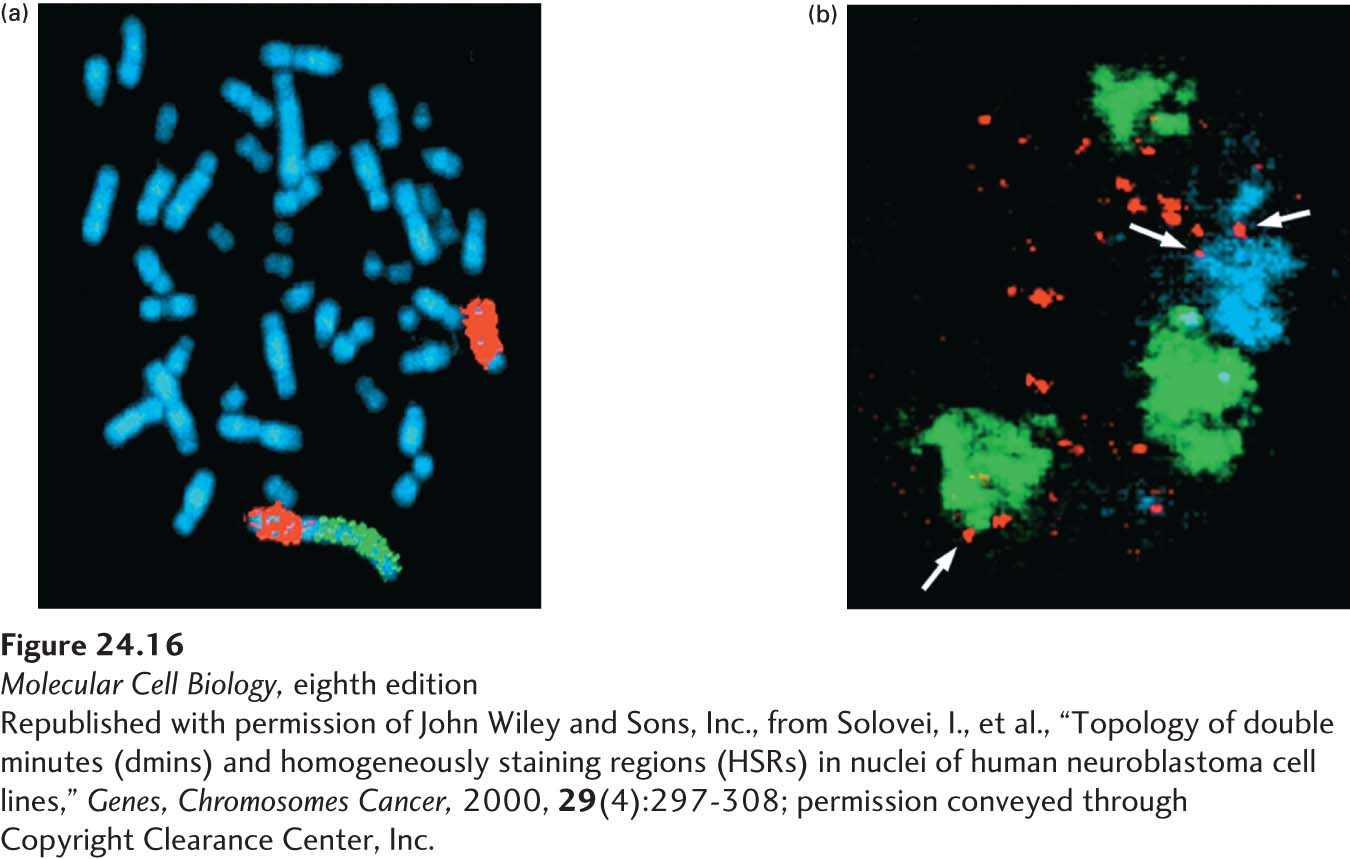
Localized amplification of DNA to produce as many as a hundred copies of a given region (usually a region spanning hundreds of kilobases) is a common genetic change seen in tumors. Normally such an event would be repaired, or the cell would be stopped from cycling by checkpoint pathways, so such lesions imply a DNA-repair defect of some kind. These amplifications may take either of two forms: the duplicated DNA may occur at a single site on a chromosome, or it may exist as small, independent mini-chromosome-like structures. The first form leads to a homogeneously staining region (HSR) that is visible in the light microscope at the site of the amplification; the second form causes extra “minute” chromosomes, separate from the normal chromosomes, that pepper a stained chromosomal preparation (Figure 24-16).
[Republished with permission of John Wiley and Sons, Inc., from Solovei, I., et al., “Topology of double minutes (dmins) and homogeneously staining regions (HSRs) in nuclei of human neuroblastoma cell lines, “Genes, Chromosomes Cancer, 2000, 29(4):297-308; permission conveyed through Copyright Clearance Center, Inc.]
EXPERIMENTAL FIGURE 24-16DNA amplifications in stained chromosomes take two forms, both visible under the light microscope. (a) Homogeneously staining regions (HSRs) in a human chromosome from a neuroblastoma cell. The chromosomes are uniformly stained with a blue dye so that all can be seen. Specific DNA sequences were detected using fluorescent in situ hybridization (FISH), in which fluorescently labeled DNA clones are hybridized to denatured DNA in the chromosomes. The chromosome 4 pair is marked by in situ hybridization with a chromosome paint probe for the long arm of chromosome 4 (red). On one of the chromosome 4’s an HSR is visible after hybridizing with a probe for the N-MYC gene (green), which is amplified in this neuroblastoma cell. (b) Optical sections through nucleus from a human neuroblastoma cell that contain so-called double-minute chromosomes. The normal chromosomes are the green and blue structures; the double-minute chromosomes are the many small red dots. Arrows indicate double minutes associated with the surface or interior of the normal chromosomes.
[Republished with permission of John Wiley and Sons, Inc., from Solovei, I., et al., “Topology of double minutes (dmins) and homogeneously staining regions (HSRs) in nuclei of human neuroblastoma cell lines, “Genes, Chromosomes Cancer, 2000, 29(4):297-308; permission conveyed through Copyright Clearance Center, Inc.]
However they arise, a central aspect of oncogenes is that the gain-of-function mutations that convert proto-oncogenes to oncogenes are genetically dominant; that is, mutation in only one of the two alleles is sufficient for induction of cancer.