Noncovalent Binding Permits Allosteric, or Cooperative, Regulation of Proteins
In addition to regulating the amount of a protein, cells can also regulate the intrinsic activity of a protein. One of the most important mechanisms for regulating protein function is through allosteric interactions. Broadly speaking, allostery (from the Greek, “other shape”) refers to any change in a protein’s tertiary or quaternary structure, or in both, induced by the noncovalent binding of a ligand. When a ligand binds to one site (A) in a protein and induces a conformational change that alters the activity of a different site (B), the ligand is called an allosteric effector of the protein, while site A is called an allosteric binding site, and the protein is called an allosteric protein. By definition, allosteric proteins have multiple binding sites, at least one for the allosteric effector and at least one for other molecules with which the protein interacts. The allosteric change in activity can be positive or negative; that is, it can be an increase or a decrease in protein activity. Negative allostery often involves the end product of a multistep biochemical pathway binding to, and reducing the activity of, an enzyme that catalyzes an early, rate-controlling step in that pathway. In this way, excessive buildup of the product is prevented. This kind of regulation of a metabolic pathway is also called end-product inhibition or feedback inhibition. Allosteric regulation is particularly prevalent in multimeric enzymes and other proteins in which conformational changes in one subunit are transmitted to an adjacent subunit.
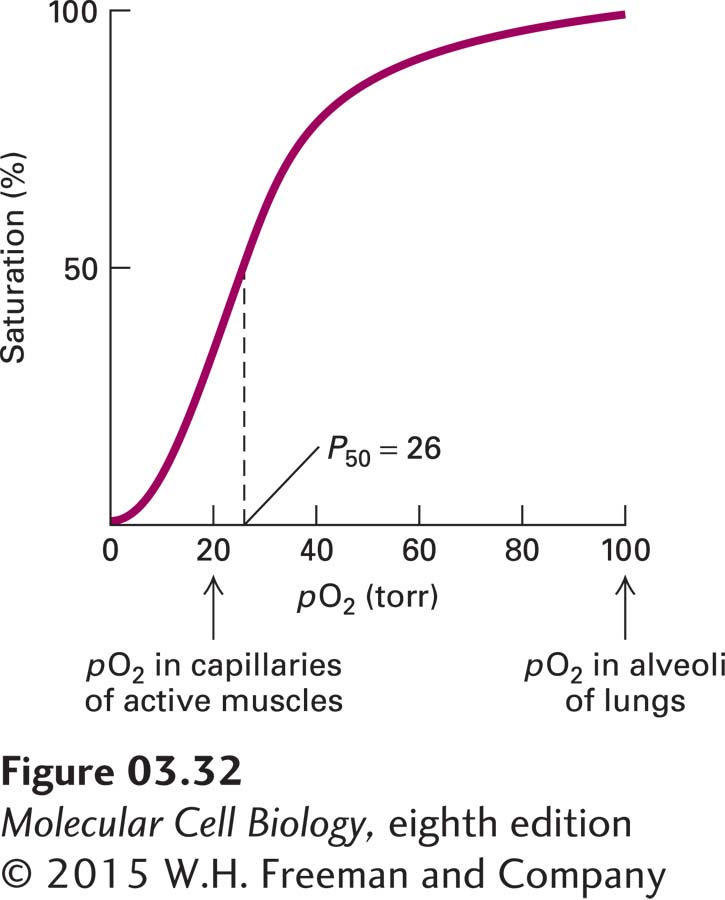
EXPERIMENTAL FIGURE 3-32Hemoglobin binds oxygen cooperatively. Each tetrameric hemoglobin molecule has four oxygen-binding sites; at saturation, all the sites are loaded with oxygen. The oxygen concentration in tissues is commonly measured as the partial pressure of oxygen (pO2) in torr units (a standard measure of pressure equivalent to 1 mm of mercury under standard conditions). P50 is the pO2 at which half the oxygen-binding sites are occupied; it is somewhat analogous to the Km for an enzymatic reaction. The large change in the amount of oxygen bound over a small range of pO2 values permits efficient unloading of oxygen in peripheral tissues such as muscle. The sigmoidal shape of a plot of saturation versus ligand concentration is indicative of cooperative binding, in which the binding of one oxygen molecule allosterically influences the binding of subsequent oxygens. In the absence of cooperative binding, a binding curve is a hyperbola, similar to the curves in Figure 3-24. See J. M. Berg et al., 2015, Biochemistry, 8th ed., Macmillan.
Cooperativity, a term that is often used synonymously with allostery, usually refers to the influence (positive or negative) that the binding of a ligand at one site has on the binding of another molecule of the same type of ligand at a different site. Hemoglobin presents a classic example of positive cooperative binding in that the binding of a single ligand, molecular oxygen (O2), increases the affinity of hemoglobin for the next oxygen molecule. Each of the four subunits in hemoglobin contains one heme molecule. The heme groups are the oxygen-binding components of hemoglobin (see Figure 3-14a). The binding of oxygen to the heme molecule in one of the four hemoglobin subunits induces a local conformational change whose effect spreads to the other subunits, lowering the Kd (increasing the affinity) for the binding of additional oxygen molecules to the remaining hemes and yielding a sigmoidal oxygen-binding curve (Figure 3-32). Because of the sigmoidal shape of the oxygen-saturation curve, it takes only a fourfold increase in oxygen concentration for the saturation of the oxygen-binding sites in hemoglobin to go from 10 to 90 percent. Conversely, if there were no cooperativity and the shape of the curve was typical of that for Michaelis-Menten (see Figure 3-24), or noncooperative, binding, it would take an eighty-one-fold increase in oxygen concentration to accomplish the same increase in loading of its binding sites in hemoglobin. This cooperativity permits hemoglobin to take up oxygen very efficiently in the lungs, where the oxygen concentration is high, and unload it in tissues, where the concentration is low. Thus cooperativity amplifies the sensitivity of a system to changes in the concentration of its ligands, providing in many cases a selective evolutionary advantage.